
Reasons for Success
It is certainly no accident that insects are the most abundant and most diverse group of organisms on earth. They have maintained a position of ecological pre-eminence for over 400 million years: they have witnessed the rise and fall of dinosaurs; they have survived at least four major cataclysms that resulted in planet-wide extinctions; and they continue to thrive despite mankind’s best efforts at eradication. While no single ecological or physiological attribute can account for this unparalleled success, the insects do have a unique combination of characteristics which, as a whole, have given them an unusual survival advantage. In brief, these attributes include an exoskeleton, small body size, the ability to fly, a high reproductive potential, complete metamorphosis, and adaptability in an ever-changing environment.
Exoskeleton
 Unlike vertebrates, an insect’s supporting skeleton is located on the outside of its body. This exoskeleton is a marvelous structure that not only gives shape and support to the body’s soft tissues, but also provides protection from attack or injury, minimizes the loss of body fluids in both arid and freshwater environments, and assures mechanical advantage to muscles for strength and agility in movement. As a “suit of armor”, the exoskeleton can resist both physical and chemical attack. It is covered by an impervious layer of wax that prevents desiccation. Much of the exoskeleton is fabricated from chitin, a polysaccharide that binds with various protein molecules to form a body wall that may be as flexible and elastic as rubber or as hard and rigid as some metals. Freedom of movement is ensured by membranes and joints in the exoskeleton. Muscles that attach directly to the body wall combine maximum strength with near-optimum mechanical advantage (leverage). The result is an ant, for example, that can lift up to 50 times its own body weight.
Unlike vertebrates, an insect’s supporting skeleton is located on the outside of its body. This exoskeleton is a marvelous structure that not only gives shape and support to the body’s soft tissues, but also provides protection from attack or injury, minimizes the loss of body fluids in both arid and freshwater environments, and assures mechanical advantage to muscles for strength and agility in movement. As a “suit of armor”, the exoskeleton can resist both physical and chemical attack. It is covered by an impervious layer of wax that prevents desiccation. Much of the exoskeleton is fabricated from chitin, a polysaccharide that binds with various protein molecules to form a body wall that may be as flexible and elastic as rubber or as hard and rigid as some metals. Freedom of movement is ensured by membranes and joints in the exoskeleton. Muscles that attach directly to the body wall combine maximum strength with near-optimum mechanical advantage (leverage). The result is an ant, for example, that can lift up to 50 times its own body weight.
Small Size
 In general, the insects are marvels of miniaturization. World’s
In general, the insects are marvels of miniaturization. World’s
Smallest
Insect Most species are between 2 and 20 mm (0.1 – 1.0 inch) in length, although they range in size from giant moths that would nearly cover your computer screen to tiny parasitic wasps that could hide inside the period at the end of this sentence.
Dichomorpha echmepterygis is the smallest of the small. Discovered in 1997, this Costa Rican wasp (family Mymaridae) is a parasite of other insects’ eggs. Adult males may be only 0.139 mm (0.00055 inch) in length — nearly 1/3 smaller than some single-celled protozoa (e.g., Paramecium caudatum).

Trail of a leaf miner larva
For an animal with an exoskeleton, small size is a distinct advantage. If insects were as large as cows or elephants, their exoskeleton would have to be proportionately thicker to support the additional mass of body tissue. But a thicker exoskeleton would also be heavier and more cumbersome. Even the simplest movements would require a larger muscle volume and consume more energy. Since a two-fold increase in body length typically results in a four-fold increase in surface area and an eight-fold increase in volume and mass, there is an upper limit to how large insects can become (somewhere around 125-150 grams). Beyond this size, the insect’s surface area is just too small for attachment of all the additional muscle tissue. “Mothra” and “The Fly” may be entertaining on the movie screen, but in real life they’d never get off the ground!!
Another advantage of small size is the minimal resources needed for survival and reproduction. A crumb is a feast; a dewdrop quenches thirst; a pebble provides shade. In some cases, food requirements are so modest that an insect may live on a single plant or animal for its entire life and never exhaust its food supply. A leaf miner, for example, spends its entire larval stage tunneling within the paper-thin layer of cells (parenchyma) between the upper and lower epidermis of a single leaf. In some ant species, an entire colony may live inside a single acorn or plant gall. Parasitic wasps in at least seven families of Hymenoptera complete their entire development within the eggs of other insects.
World’s
Biggest
InsectsFinally, small size is a big advantage to insects that must avoid predation. They can hide in the cracks of a rock, beneath the bark of a tree, behind the petal of a flower, or under a blade of grass. The exoskeleton is hard enough for them to burrow between individual grains of sand, yet flexible enough to let them squeeze through the tiniest of cracks. Small size, together with adaptations in body shape and coloration, gives many species the ability to blend so well with their environment that they become virtually undetectable.
Flight
 Insects are the only invertebrates that can fly. Judging from the fossil record, they acquired this ability about 300 million years ago — nearly 100 million years before the advent of the first flying reptiles. Flight gave these insects a highly effective mode of escape from predators that roamed the prehistoric landscape. It was also an efficient means of transportation, allowing populations to expand more quickly into new habitats and exploit new resources.
Insects are the only invertebrates that can fly. Judging from the fossil record, they acquired this ability about 300 million years ago — nearly 100 million years before the advent of the first flying reptiles. Flight gave these insects a highly effective mode of escape from predators that roamed the prehistoric landscape. It was also an efficient means of transportation, allowing populations to expand more quickly into new habitats and exploit new resources.
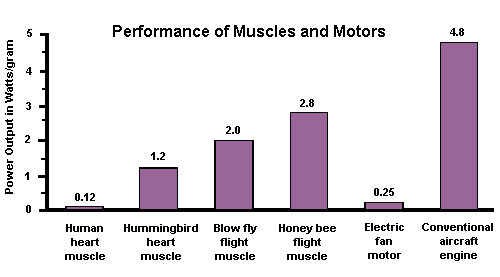
The forces of natural selection have shaped insect flight into a remarkably efficient process employing principles of aerodynamics that human engineers do not yet fully understand. Although the metabolic cost of flight (calories per unit of lift) is similar to that of birds and bats, an insect’s flight musculature produces at least 2X more power per unit of muscle mass (see chart below). This high efficiency is largely due to elasticity of the thorax — 90-95% of the potential energy absorbed by flexion of the exoskeleton is released as kinetic energy during the wing’s downstroke.
Efficient use of energy allows some insects to travel great distances or remain airborne for long periods of time. More than 200 species, including moths, dragonflies, locusts, flies, and beetles are known to migrate over long distances by air. The migratory locust, Schistocerca gregaria, can fly for up to 9 hours without stopping. Large swarms occasionally traverse the Mediterranean Sea. In North America, annual migrations of monarch butterflies (Danaus plexippus) wing their way from summer feeding grounds to overwintering sites in California and Mexico. One individual, marked in Ontario, Canada was collected four months later in Mexico, 2800 km from its birthplace.
 Speed and maneuverability are traits usually associated with the more highly advanced orders of insects, but some primitive insects (notably the dragonflies) are remarkably agile fliers: they can hover, turn while hovering, and even fly backwards or sideways. For their size, some species are remarkably swift. Large hawker dragonflies (family Ashnidae) have been clocked at a top speed of 58 km/hr (36 mph) over level ground. By comparison, the fastest human sprinters run only about 36 km/hr (22.5 mph).
Speed and maneuverability are traits usually associated with the more highly advanced orders of insects, but some primitive insects (notably the dragonflies) are remarkably agile fliers: they can hover, turn while hovering, and even fly backwards or sideways. For their size, some species are remarkably swift. Large hawker dragonflies (family Ashnidae) have been clocked at a top speed of 58 km/hr (36 mph) over level ground. By comparison, the fastest human sprinters run only about 36 km/hr (22.5 mph).
The wings of a large insect can generate a considerable amount of lift. Green darner dragonflies (Anax junius) are able to fly while carrying a load up to 15 times their body weight. Insects with smaller wings have to work much harder to remain airborne. Some of the tiny biting midges (like Forcipomyia spp.) beat their wings over 1000 times per second. A special type of muscle tissue is needed to sustain this rapid rate of contraction.
Reproductive Potential
 Reproductive success is one of the most significant measures of an organism’s fitness. In insect populations, females often produce large numbers of eggs (high fecundity), most of the eggs hatch (high fertility), and the life cycle is relatively short (often as little as 2-4 weeks). Together, these three characteristics enable insects to produce remarkably large numbers of offspring. A typical female lays 100-500 eggs in her lifetime, but numbers in the thousands are not uncommon. The queen of an African termite colony may be the mother of more than ten million workers during her 20-25 year lifespan.
Reproductive success is one of the most significant measures of an organism’s fitness. In insect populations, females often produce large numbers of eggs (high fecundity), most of the eggs hatch (high fertility), and the life cycle is relatively short (often as little as 2-4 weeks). Together, these three characteristics enable insects to produce remarkably large numbers of offspring. A typical female lays 100-500 eggs in her lifetime, but numbers in the thousands are not uncommon. The queen of an African termite colony may be the mother of more than ten million workers during her 20-25 year lifespan.
Antoni van Leeuwenhoek, the Dutch scientist who first discovered single-celled organisms, was probably the first person to comprehend the reproductive potential of insects. In 1687, he reared blow flies (probably Calliphora erythrocephala) on a diet of owl meat and found that females produced an average of 144 progeny: half male, half female. Using this data (and assuming no mortality), he calculated that a single pair of flies could give rise to 10,368 offspring in the third generation, 746,496 in the fourth generation, 53,747,712 in the fifth generation, and 3,869,835,264 in the sixth generation. At this rate, the fly population would grow to more than 1 X 1017 individuals in one year (10 generations). In two years of continuous reproduction the flies would fill a ball nearly 8 million kilometers in diameter. Calculations like this are completely unrealistic because they ignore all the limiting factors (like food supply, predation, climate, and disease) that keep natural populations in check. But the numbers do illustrate how quickly insect populations can grow, and they help explain why pest outbreaks often seem to develop so suddenly.
Since most insects die before they ever have an opportunity to reproduce, a high reproductive potential is the species’ best chance for survival. Many adaptations help maximize this potential. Most females, for example, can store sperm for months or years within the spermatheca, a special region of the reproductive system. A single mating can supply a female with enough sperm to fertilize all the eggs she will produce in her lifetime. An unbalanced sex ratio, where females outnumber males, is another way to maximize reproductive potential. Since most insects are not monogamous, a few males can supply sperm for a large number of females. And finally, there are many species (e.g. aphids, scale insects, thrips, and midges) where males are entirely absent — all members of the population are female and contribute offspring through a process of asexual reproduction.
Metamorphosis

Life cycle of the house fly
Most insects undergo significant developmental changes as they grow from immatures to adults. These changes, collectively known as metamorphosis, may involve physical, biochemical, and/or behavioral alterations that promote survival, dispersal, and reproduction of the species. In the more primitive insects, most of these changes occur gradually as the animal matures: organs of reproduction and flight develop incrementally during the immature stages and become functional only in adults. Since this transformation process is slow and does not include all body tissues (incomplete metamorphosis) the immatures and adults share many characteristics — they often live in similar habitats and feed on similar types of food.
More advanced insects, however, undergo complete metamorphosis — a dramatic transformation in form and function between the immature (larval) and adult stages of development. In these insects, a larva is primarily adapted for feeding and growth. It assimilates energy reserves which, in some cases, will sustain the insect for the rest of its life. When fully grown, a larva molts into a transitional stage, called the pupa, and begins a period of massive internal and external reorganization. Body organs and tissues encoded by larval DNA are disassembled and rebuilt according to a second DNA blueprint that had been repressed during larval life. An adult insect (imago) eventually emerges from within the pupal exoskeleton bearing little or no resemblance to its larval form. Its primary function is dispersal and reproduction.
In the class Insecta, only 9 out of 28 orders undergo complete metamorphosis, yet these 9 orders represent about 86% of all insect species alive today. The obvious advantage to this type of development lies in the compartmentalization of the life cycle. Through natural selection, larval form and function can be optimized for growth and feeding without compromising adaptations of the adult for dispersal and reproduction. Each stage of the life cycle is entirely free to adapt to its own ecological role. In some cases, this means that immatures and adults may consume different types of food, exploit different environmental resources, and even occupy different habitats.
Adaptability
 A combination of large and diverse populations, high reproductive potential, and relatively short life cycles, has equipped most insects with the genetic resources to adapt quickly in the face of a changing environment. Their record of achievement is impressive: they were among the first creatures to invade the arid expanses of dry land and exploit green plants as a source of food, they were the first animals to use flight as an escape from predators, and they were the first organisms to develop a complex social hierarchy with division of labor and cooperative care of the young. As a group, they have endured 400 million years of climatological and geophysical upheaval, including the evaporation of inland seas, formation of mountain ranges, shifts in continental plates, onset of ice ages, and the fallout from cosmic impacts.
A combination of large and diverse populations, high reproductive potential, and relatively short life cycles, has equipped most insects with the genetic resources to adapt quickly in the face of a changing environment. Their record of achievement is impressive: they were among the first creatures to invade the arid expanses of dry land and exploit green plants as a source of food, they were the first animals to use flight as an escape from predators, and they were the first organisms to develop a complex social hierarchy with division of labor and cooperative care of the young. As a group, they have endured 400 million years of climatological and geophysical upheaval, including the evaporation of inland seas, formation of mountain ranges, shifts in continental plates, onset of ice ages, and the fallout from cosmic impacts.
Adaptation is an ongoing process. Populations must continually change as new resources appear and old ones disappear. Just within the few thousand years since humans began roaming the earth, insects have acquired a taste for new products that would never be a part of their “natural” environment: e.g., glue and wallpaper paste, book bindings, cardboard, paintbrushes, tanned leather products, the corks of wine bottles, mummies, stuffed museum specimens, chocolate, ginger, yeast cakes, tobacco, pepper, and even potent drugs like marijuana and opium. In the early 1900’s, a powder post beetle (Scobicia declivis) made itself a nuisance by gnawing holes through the lead sheath around telephone and telegraph cables. Once inside the cable, these “short-circuit beetles” ate the fiber insulator around the copper conductor. After enough insulation was gone, the wire would touch the lead sheath and create a short-circuit in the line. The high cost of replacing damaged cable eventually forced the telephone company to develop non-fibrous insulators for their long-distance cables.
 Perhaps the most remarkable example of insect adaptation in this century has been the speed with which pest populations have developed resistance to a broad range of chemical and biological insecticides. After World War II, public health officials in the United States made a concerted effort to eradicate the house fly (Musca domestica) with DDT. For several years the campaign seemed promising: fly populations decreased and optimism ran high. But a few resistant flies managed to survive because they were endowed with an enzyme that could detoxify DDT. These survivors reproduced and passed this resistant trait to their offspring. In time, DDT-resistant flies repopulated their environment and the species now appears to be living happily ever after!
Perhaps the most remarkable example of insect adaptation in this century has been the speed with which pest populations have developed resistance to a broad range of chemical and biological insecticides. After World War II, public health officials in the United States made a concerted effort to eradicate the house fly (Musca domestica) with DDT. For several years the campaign seemed promising: fly populations decreased and optimism ran high. But a few resistant flies managed to survive because they were endowed with an enzyme that could detoxify DDT. These survivors reproduced and passed this resistant trait to their offspring. In time, DDT-resistant flies repopulated their environment and the species now appears to be living happily ever after!
Significant levels of pesticide resistence have now been reported in over 500 insect species, and many of these animals are resistant to compounds from more than one chemical family. In New York, for example, potato growers on Long Island must cope with a population of the Colorado potato beetle (Leptinotarsa decemlineata) that is resistant not only to organophosphates, carbamates, and synthetic pyrethroids, but also to some of the insect growth regulators and microbial insecticides.
