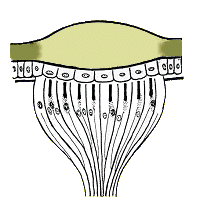
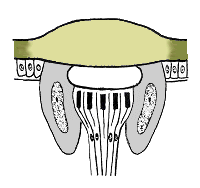
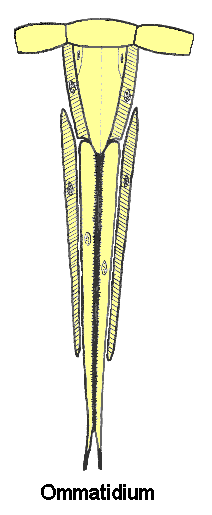
 The light−sensitive part of an ommatidium is called the rhabdom. It is a rod-like structure, secreted by an array of 6-8 specialized neurons (retinula cells), and centered on the optical axis just below the crystalline cone. The rhabdom contains an array of closely packed microtubules where light-sensitive pigments (e.g. rhodopsin, etc.) are stored. These pigments absorb certain wavelengths of incident light and generate nerve impulses through a photochemical process similar to that of vertebrates.
The light−sensitive part of an ommatidium is called the rhabdom. It is a rod-like structure, secreted by an array of 6-8 specialized neurons (retinula cells), and centered on the optical axis just below the crystalline cone. The rhabdom contains an array of closely packed microtubules where light-sensitive pigments (e.g. rhodopsin, etc.) are stored. These pigments absorb certain wavelengths of incident light and generate nerve impulses through a photochemical process similar to that of vertebrates.
Most diurnal insects have pigment cells surrounding each ommatidium. These cells limit a facet’s field of view by absorbing light that enters through adjacent corneas. Each facet points toward a slightly different part of the visual field. In composite, they render a mosaic-like impression of the environment. Nocturnal and crepuscular insects have pigment cells that do not completely isolate each facet. Their ommatidia are stimulated by light from larger fields of view. This produces a brighter but theoretically less distinct mosaic image.
Since insects cannot form a true (i.e. focused) image of the environment, their visual acuity is relatively poor compared to that of vertebrates. On the other hand, their ability to sense movement, by tracking objects from ommatidium to ommatidium, is superior to most other animals. Temporal resolution of flicker is as high as as 200 images/second in some bees and flies (in humans, still images blur into constant motion at about 30 images/second).
Unlike humans, most insects can distinguish between polarized light (coming directly from the sun) and unpolarized light (reflected from water vapor and other particles in the atmosphere). This ability allows them to detect the sun’s position in the sky, even on cloudy or overcast days, and use it as an orientation cue.
Compared to humans, insects have a range of spectral sensitivity that is shifted toward shorter wavelengths (higher frequencies). Thus, insects can “see” light in the ultraviolet range that is invisible to humans. On the other hand, insects cannot detect wavelengths at the red end of the spectrum that are visible to humans. True color vision, however, involves more than just a wide range of spectral sensitivity. Most insects have only a limited ability to discriminate different colors of light, but a few (especially bees and butterflies) have “true” color vision.

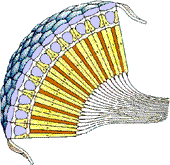 A pair of compound eyes are the principle visual organs of most insects; they are found in nearly all adults and in many immatures of ametabolous and hemimetabolous orders. As the name suggests, compound eyes are composed of many similar, closely-packed facets (called ommatidia) which are the structural and functional units of vision. The number of ommatidia varies considerably from species to species: some worker ants have fewer than six while some dragonflies may have more than 25,000.
A pair of compound eyes are the principle visual organs of most insects; they are found in nearly all adults and in many immatures of ametabolous and hemimetabolous orders. As the name suggests, compound eyes are composed of many similar, closely-packed facets (called ommatidia) which are the structural and functional units of vision. The number of ommatidia varies considerably from species to species: some worker ants have fewer than six while some dragonflies may have more than 25,000.