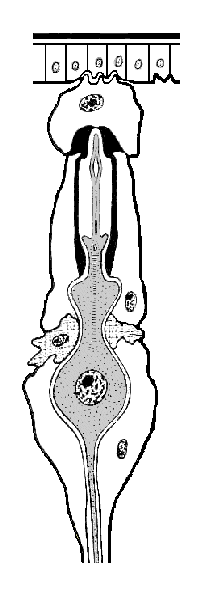
Mechanoreceptors
Insect mechanoreceptors can be found almost anywhere on the surface of an insect’s body. They may act as tactile receptors, detecting movement of objects in the environment, or they may provide proprioceptive cues (sensory input about the position or orientation of the body and its appendages). These receptors are innervated by one or more sensory neurons that fire in response to stretching, bending, compression, vibration, or other mechanical disturbance. Some mechanoreceptors produce a phasic response when stimulated — that is, they fire once when activated and again when deactivated. Other receptors generate a tonic response, firing repeatedly as long as a stimulus persists. Neural processing centers in the brain or segmental ganglia interpret the combinations of tonic and phasic signals sent from nearby receptors.
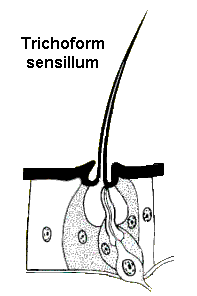 Trichoform sensilla are probably the simplest mechanoreceptors. These are tactile hairs (setae) that are innervated by a sensory neuron. Dendrites of the neuron attach near the base of the hair and generate a nerve impulse whenever they detect movement. Hair beds (clusters of tactile setae) are often found behind the head, on the legs, or near joints where they respond to movements of the body.
Trichoform sensilla are probably the simplest mechanoreceptors. These are tactile hairs (setae) that are innervated by a sensory neuron. Dendrites of the neuron attach near the base of the hair and generate a nerve impulse whenever they detect movement. Hair beds (clusters of tactile setae) are often found behind the head, on the legs, or near joints where they respond to movements of the body.
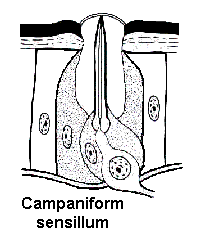 Campaniform sensilla are flattened oval discs that usually serve as flex receptors in the exoskeleton. They respond whenever mechanical stress causes the exoskeleton to bend. Campaniform sensilla are found throughout the body — especially on the legs, near the base of the wings, and along sutures where two sclerites of the exoskeleton meet.
Campaniform sensilla are flattened oval discs that usually serve as flex receptors in the exoskeleton. They respond whenever mechanical stress causes the exoskeleton to bend. Campaniform sensilla are found throughout the body — especially on the legs, near the base of the wings, and along sutures where two sclerites of the exoskeleton meet.
Stretch receptors are multi-polar neurons that usually accompany muscle or connective tissue. They are commonly embedded in intersegmental membranes and in the muscular walls of the digestive system. Some insects may stop feeding when the gut expands enough to stimulate stretch receptors in the crop. In others, oviposition may begin when mature eggs stimulate stretch receptors in the reproductive system.
Pressure receptors provide sensory information about an aquatic insect’s depth in the water. These receptors are usually associated with a cushion of air against the body or within the tracheal system. Increasing water pressure deflects hair-like processes within the receptor and stimulates tonic and phasic impulses.