
Morphogenesis
Once an insect hatches from the egg it is usually able to survive on its own, but it is small, wingless, and sexually immature. Its primary role in life is to eat and grow. If it survives, it will periodically outgrow and replace its exoskeleton (a process known as molting). In many species, there are other physical changes that also occur as the insect gets older (growth of wings and development of external genitalia, for example). Collectively, all changes that involve growth, molting, and maturation are known as morphogenesis.
Molting
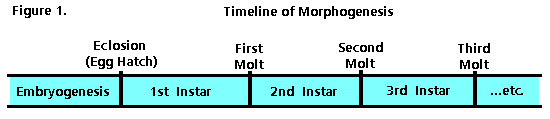
The molting process is triggered by hormones released when an insect’s growth reaches the physical limits of its exoskeleton. Each molt represents the end of one growth stage (instar) and the beginning of another (Figure 1). In some insect species the number of instars is constant (typically from 3 to 15), but in others it may vary in response to temperature, food availability, or other environmental factors. An insect is known as an imago (adult) when it becomes sexually mature. At this point, molting stops and energy for growth is channeled into production of eggs or sperm.
An insect cannot survive without the support and protection of its exoskeleton, so a new, larger replacement must be constructed inside the old one — much like putting an overcoat under a sweater! The molting process begins when epidermal cells respond to hormonal changes by increasing their rate of protein synthesis. This quickly leads to apolysis — physical separation of the epidermis from the old endocuticle. Epidermal cells fill the resulting gap with an inactive molting fluid and then secrete a special lipoprotein (the cuticulin layer) that insulates and protects them from the molting fluid’s digestive action. This cuticulin layer becomes part of the new exoskeleton’s epicuticle.











