
Holometabola: The Endopterygote Group
Complete metamorphosis (holometabolous development), is the most distinctive characteristic of all endopterygote orders. These insects have four developmental stages in the life cycle: egg, larva, pupa, and adult (imago). The larval stage is a period of active feeding and growth. The pupal stage is a period of reconstruction: larval tissues are broken down (histolyzed) and rebuilt according to the adult body plan. The adult stage is a period of dispersal and reproduction.
The ancestors of today’s holometabolous insects may have first appeared near the end of the Carboniferous period (around 290 million years ago) — probably not long after Hemipteroids and Orthopteroids diverged from their ancestral forms. From an evolutionary point of view, this event is nearly as significant as the development of wings because it marks the point at which adaptions of immatures (for feeding and growth) became functionally independent from adaptations of the adults (for reproduction and dispersal). Unlike hemimetabolous insects in which the immature structures (e.g., legs, eyes, antennae, etc.) must also serve the adults, holometabolous insects acquire a completely new body during the pupal stage. In effect, the insect is pre-programmed with two sets of genetic instructions, one for the larval stage and a different one for the adult stage. This complete transformation makes it possible for larvae and adults to respond to selective pressures in different ways, to develop independent adaptations, and even to evolve very different lifestyles.
The selective advantage of holometabolous development is under-scored by today’s dominance of these insects in the world’s fauna. Although endopterygote orders represent only about 1/3 of the living insect orders, they contain over 4/5 of the known insect species. In fact, the four largest orders of insects, based on number of species, are all members of the Holometabola. Mother Nature likes good breeding!
Despite a reasonably good fossil record, it is still impossible to trace any endopterygote lineage all the way back to its Permian ancestors. Consequently, proposed phylogenetic relationships among the Holometabola remain largely speculative. Our current theories are “educated guesses”; they are only as good as the assumptions upon which they are based.
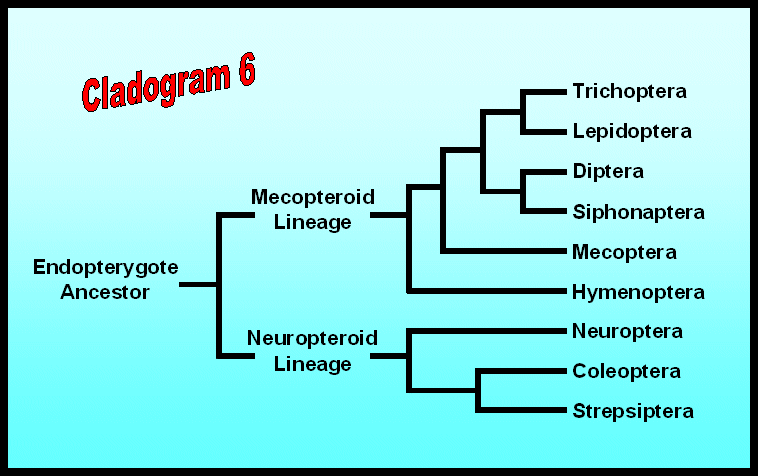
Most taxonomists assume that the first holometabolous insects were relatively “unspecialized”: mandibulate mouthparts, four similar wings with heavy venation, five-segmented tarsi, and active immature stages. Because Mecoptera (scorpionflies) and Neuroptera (lacewings and their relatives) are the two endopterygote orders that most nearly fit this description, either one might represent the ancestral body plan. In fact, both orders may have a legitimate claim to this distinction because fossils of each order are among the first to appear in rocks of the Permian period. It is certainly reasonable to suggest that two sister groups diverged from a primitive endopterygote ancestor and that all living members of the Holometabola descended from one of these two lineages (Cladogram 6). If this interpretation is not correct, then there may be as many as six separate lines of evolution leading away from a common endopterygote ancestor.
One noticable trend in the phylogeny of endopterygote orders is a structural or functional reduction in the number of wings. The most primitive orders have two pairs of wings, all more or less similar in size, with independent musculature and asynchronous wing beat. In the more specialized orders, one or more pair of wings becomes vestigial (Siphonaptera), modified in function (Diptera, Coleoptera, and Strepsiptera), or coupled together with other wings by means of hooks, hairs, or bristles to act as a single flight surface (Hymenoptera, Trichoptera, and Lepidoptera).
Neuroptera
 The order Neuroptera includes the lacewings and antlions. All members of the order have grasping/biting mouthparts and the larval stages exhibit a predatory lifestyle. Except for larval spongillaflies (family Sisyridae) which feed on fresh-water sponges, all members are terrestrial. Antlion larvae live in the soil and construct pitfall traps to snare prey. Lacewing larvae are usually found in vegetation where they typically feed on aphids, mites, and scale insects. In most cases, the adults of these insects are also predators — the non-predatory species usually feed on nectar, pollen, or honeydew.
The order Neuroptera includes the lacewings and antlions. All members of the order have grasping/biting mouthparts and the larval stages exhibit a predatory lifestyle. Except for larval spongillaflies (family Sisyridae) which feed on fresh-water sponges, all members are terrestrial. Antlion larvae live in the soil and construct pitfall traps to snare prey. Lacewing larvae are usually found in vegetation where they typically feed on aphids, mites, and scale insects. In most cases, the adults of these insects are also predators — the non-predatory species usually feed on nectar, pollen, or honeydew.
The larvae of antlions and lacewings have specialized mouthparts with large, sickle-shaped mandibles and maxillae that interlock to form pincers. Once impaled on these pincers, a prey’s body contents are sucked out through hollow food channels running between the adjacent surfaces of the mandibles and maxillae.
As adults, all neuropterans have two pairs of membranous wings with an extensive pattern of veins and crossveins. At rest, the wings are folded flat over the abdomen or held tent-like over the body. Most species are rather weak fliers.
When they pupate, larvae of lacewings and antlions More about
Neuroptera dig a small cavity in the soil and spin a loose silken cocoon around themselves. Many holometabolous insects exhibit similar behavior, but neuropterans are unusual because their silk is produced by Malpighian tubules (excretory organs) and spun from the anus. In contrast, most other endopterygote insects produce silk in modified salivary glands and spin it with mouthparts. Only one other order, the Coleoptera, makes silk in the same manner as Neuroptera.
Megaloptera
 The order Megaloptera includes the dobsonflies and alderflies. The Megaloptera are always aquatic as immatures. They live under stones or submerged vegetation and feed on a variety of small aquatic organisms. More about
The order Megaloptera includes the dobsonflies and alderflies. The Megaloptera are always aquatic as immatures. They live under stones or submerged vegetation and feed on a variety of small aquatic organisms. More about
Megaloptera Large species, often called hellgrammites, may require several years of growth to reach maturity. Adults usually remain near water, although they are attracted to lights at night. In most species, the adults live only a few days and rarely feed. In the past, the Megaloptera were often classified as a suborder of Neuroptera.
Raphidioptera
 The order Raphidioptera includes the snakeflies which are terrestrial. Snakefly larvae live in leaf litter or under bark and catch aphids or other soft-bodied prey. More about
The order Raphidioptera includes the snakeflies which are terrestrial. Snakefly larvae live in leaf litter or under bark and catch aphids or other soft-bodied prey. More about
Raphidioptera In most cases, the adults of these insects are also predators — the non-predatory species usually feed on nectar, pollen, or honeydew. In the past, the Raphidioptera were often classified as a suborder of Neuroptera.
Coleoptera
 A phylogenetic link between Neuroptera and Coleoptera is further strengthened by the existence of Permian fossils that appear to share the characteristics of both orders. Some of these beetle-like insects evidently evolved as carnivores (suborder Aedephaga) while others were herbivores (suborder Polyphaga). Significant growth and diversification of the herbivore lineage occurred near the beginning of the Cretaceous period (100-130 million years ago) when flowering plants (angiosperms) first began to take over as the dominant form of plant life.
A phylogenetic link between Neuroptera and Coleoptera is further strengthened by the existence of Permian fossils that appear to share the characteristics of both orders. Some of these beetle-like insects evidently evolved as carnivores (suborder Aedephaga) while others were herbivores (suborder Polyphaga). Significant growth and diversification of the herbivore lineage occurred near the beginning of the Cretaceous period (100-130 million years ago) when flowering plants (angiosperms) first began to take over as the dominant form of plant life.
Coleoptera (beetles and weevils) is the largest order in the class Insecta. As adults, most beetles have a hard, dense exoskeleton that covers and protects most of their body surface. The front wings, known as elytra, are just as hard as the rest of the exoskeleton. They fold down over the abdomen and serve as protective covers for the large, membranous hind wings. At rest, both elytra meet along the middle of the back, forming a straight line that is probably the most distinctive characteristic of the order. During flight, the elytra are held out to the sides of the body where they provide a certain amount of aerodynamic stability.
Both larvae and adults have strong mandibulate mouthparts. As a group, they feed on a wide variety of diets, inhabit all terrestrial and fresh-water environments, and exhibit a number of different life-styles. Many species are herbivores — variously adapted to feed on the roots, stems, leaves, or reproductive structures of their host plants. Some species live on fungi, others burrow into plant tissues, still others excavate tunnels in wood or under bark.
Many beetles are predators. More about
Coleoptera They live in the soil or on vegetation and attack a wide variety of invertebrate hosts. Some beetles are scavengers, feeding primarily on carrion, fecal material, decaying wood, or other dead organic matter. There are even a few parasitic beetles — some are internal parasites of other insects, some invade the nests of ants or termites, and some are external parasites of mammals.
Strepsiptera
 Most beetle larvae that parasitize insects are free-living as adults. But the stylopids (order Strepsiptera) are thought to be a closely related group that have retained the parasitic lifestyle even as adults. Most Strepsiptera (also known as twisted-wing parasites) live as internal parasites of bees, wasps, grasshoppers, leafhoppers, and other members of the order Hemiptera. Only a few species that parasitize bristletails (Archeognatha) are known to be free-living in the adult stage.
Most beetle larvae that parasitize insects are free-living as adults. But the stylopids (order Strepsiptera) are thought to be a closely related group that have retained the parasitic lifestyle even as adults. Most Strepsiptera (also known as twisted-wing parasites) live as internal parasites of bees, wasps, grasshoppers, leafhoppers, and other members of the order Hemiptera. Only a few species that parasitize bristletails (Archeognatha) are known to be free-living in the adult stage.
Strepsiptera share so many characteristics with beetles that some entomologists classify them as a superfamily of Coleoptera. In fact, Strepsiptera and certain parasitic beetles (in the families Meloidae and Rhipiphoridae) are among the very few insects that undergo hypermetamorphosis, an unusual type of holometabolous development in which the larvae change body form as they mature. Upon emerging from their mother’s body, the young larvae, called triunguloids, have six legs and crawl around in search of a suitable host. In species that parasitize bees or wasps, a triunguloid usually climbs to the top of a flower and waits for a pollinator. When a host arrives, the larva burrows into its body and quickly molts into a second stage that has no distinct head, legs, antennae or other insect-like features. These larvae grow and continue to molt inside the host’s body cavity, assimilating nutrients from the blood and non-vital tissues. After pupating in the host, winged males emerge and fly in search of mates. An adult female remains inside her host, managing to attract and mate with a male while only a small portion of her body protrudes from the host’s abdomen. Embryos develop within the female’s body, and a new generation of triunguloid larvae begin their life cycle by escaping through a brood passage on the underside of her body.
Adult male Strepsiptera are strange-looking insects. More about
Strepsiptera The head is small, with protruding compound eyes that look like tiny raspberries. The antennae are multi-segmented and have up to three branches. Front wings are reduced to small, club-like structures; hind wings are very large and fan-shaped. In fact, Coleoptera and male Strepsiptera are the only holometabolous insects in which the hind wings are larger than the front wings and provide most of the lift during flight.
Mecoptera
 At the same time beetles were diverging from their neuropteran ancestors, the Mecoptera were also diverging and specializing. In this lineage, front wings became the predominate flight surfaces. The hind wings were either modified in function (Diptera) or else reduced in size and linked together with the front wings (by means of hooks, hairs, or bristles) to form a single flight surface.
At the same time beetles were diverging from their neuropteran ancestors, the Mecoptera were also diverging and specializing. In this lineage, front wings became the predominate flight surfaces. The hind wings were either modified in function (Diptera) or else reduced in size and linked together with the front wings (by means of hooks, hairs, or bristles) to form a single flight surface.
The Mecoptera (scorpionflies) are a curious group of terrestrial insects that usually live in moist sylvan habitats. Both larvae and adults are omnivorous. Mostly, they feed upon decaying vegetation and dead (or dying) insects. Larvae generally remain in the soil; they have chewing mouthparts and resemble caterpillars (Lepidoptera) or white grubs (Coleoptera). Most adults have an elongated head with slender, chewing mouthparts near the tip of a stout beak. Front and hind wings are similar in shape (occasionally reduced in size or absent), and often mottled with patches of color. The common name of this order (scorpionfly) refers to the distinctive appearance of male genitalia in members of the family Panorpidae: the terminal segments are enlarged and held recurved over the abdomen like the tail of a scorpion. Despite its appearance, the scorpionfly’s tail is quite harmless.
Hanging scorpionflies, family Bittacidae, are predators of small flying insects. More about
Mecoptera Their legs, especially the tarsi, are unusually long and slender. At the tip of each leg there is a single opposable claw. The adults hang from vegetation with their front legs and catch small flying insects with their middle and hind legs. These scorpionflies, which bear a striking resemblance to crane flies (Diptera: Tipulidae), may have developed from the same ancestral lineage that also give rise to the caddisflies (order Trichoptera) and the true flies (order Diptera).
Diptera
 The order Diptera includes all the true flies. These insects are distinctive because their hind wings are reduced to small, club-shaped structures called halteres — only the membranous front wings serve as aerodynamic surfaces. The halteres, which vibrate during flight, work much like a gyroscope to help the insect maintain balance while it is in flight.
The order Diptera includes all the true flies. These insects are distinctive because their hind wings are reduced to small, club-shaped structures called halteres — only the membranous front wings serve as aerodynamic surfaces. The halteres, which vibrate during flight, work much like a gyroscope to help the insect maintain balance while it is in flight.
All Dipteran larvae are legless. They live in aquatic (fresh water), semi-aquatic, or moist terrestrial environments. They are commonly found in the soil, in plant or animal tissues, and in carrion or dung — almost always where there is little danger of desiccation. Some species are herbivores, but most feed on dead organic matter or parasitize other animals, especially vertebrates, molluscs, and other arthropods. In the more primitive families (suborder Nematocera), fly larvae have well-developed head capsules with mandibulate mouthparts. These structures are reduced or absent in the more advance suborders (Brachycera and Cyclorrhapha) where the larvae, known as maggots, have worm-like bodies and only a pair of mouth hooks for feeding.
Adult flies live in a wide range of habitats and display enormous variation in appearance and life style. Although most species have haustellate mouthparts and collect food in liquid form, their mouthparts are so diverse that some entomologists suspect the feeding adaptations may have arisen from more than a single evolutionary origin. In many families, the proboscis (rostrum) is adapted for sponging and/or lapping.
These flies survive on honeydew, nectar, or the exudates of various plants and animals (dead or alive). More about
Diptera In other families, the proboscis is adapted for cutting or piercing the tissues of a host. Some of these flies are predators of other arthropods (e.g., robber flies), but most of them are external parasites (e.g., mosquitoes and deer flies) that feed on the blood of their vertebrate hosts, including humans and most wild and domestic animals.
Siphonaptera
 Judging from their haustellate mouthparts, Siphonaptera (fleas) may have diverged from one of the dipteran lineages. As adults, all fleas are blood-sucking external parasites. Most species feed on mammals, although a few (less than 10%) live on birds. Only adult fleas inhabit the host’s body and feed on its blood. They are active insects with a hard exoskeleton, strong hind legs adapted for jumping, and a laterally flattened body that can move easily within the host’s fur or feathers. Unlike lice, most fleas spend a considerable amount of time away from their host. Adults may live for a year or more and can survive for weeks or months without a blood meal.
Judging from their haustellate mouthparts, Siphonaptera (fleas) may have diverged from one of the dipteran lineages. As adults, all fleas are blood-sucking external parasites. Most species feed on mammals, although a few (less than 10%) live on birds. Only adult fleas inhabit the host’s body and feed on its blood. They are active insects with a hard exoskeleton, strong hind legs adapted for jumping, and a laterally flattened body that can move easily within the host’s fur or feathers. Unlike lice, most fleas spend a considerable amount of time away from their host. Adults may live for a year or more and can survive for weeks or months without a blood meal.
Flea larvae are worm-like (vermiform) in shape with a sparse covering of bristles. More about
Siphonaptera They rarely live on the body of their host. Instead, they are usually found in its nest or bedding where they feed as scavengers on organic debris (including adult feces). In general, flea larvae can survive more arid conditions than most fly larvae. After a larval period that includes two molts, fleas pupate within a thin silken cocoon. Under favorable conditions, the life cycle can be completed in less than a month.
Trichoptera
 The order Trichoptera (caddisflies) is another likely descendant of the Mecopteran lineage. Adults are mostly nocturnal, weak-flying insects that are often attracted to lights. During the day, they hide in cool, moist environments such as the vegetation along river banks. The body and wings are clothed with long silky hairs (setae) — a distinctive characteristic of the order. In flight, the hind wings are coupled to the front wings by specially curved hairs. At rest the wings are held tent-like over the abdomen. Many caddisflies have reduced or vestigal mouthparts. Few species have actually been observed feeding, and most adults are relatively short-lived.
The order Trichoptera (caddisflies) is another likely descendant of the Mecopteran lineage. Adults are mostly nocturnal, weak-flying insects that are often attracted to lights. During the day, they hide in cool, moist environments such as the vegetation along river banks. The body and wings are clothed with long silky hairs (setae) — a distinctive characteristic of the order. In flight, the hind wings are coupled to the front wings by specially curved hairs. At rest the wings are held tent-like over the abdomen. Many caddisflies have reduced or vestigal mouthparts. Few species have actually been observed feeding, and most adults are relatively short-lived.
All caddisfly larvae live in aquatic environments; More about
Trichoptera they may be herbivores, scavengers, or predators. In most cases, the predatory species are free-living or spin silken structures in the water (webs or tunnels) to entrap prey. The scavengers and herbivores live within protective “cases” which they build from their own silk and stones, twigs, leaf fragments, or other natural materials.
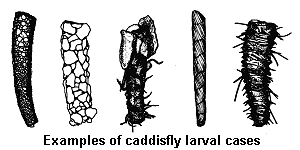 Case design and construction is distinctive for each family or genus of caddisfly. The case is usually portable, dragged around like a snail shell as the insect moves, and held in place by a pair of hooked prolegs at the tip of the abdomen. Most species have thread-like abdominal gills and get oxygen from water that circulates inside the case. All larval growth and development (including pupation) occurs within the case.
Case design and construction is distinctive for each family or genus of caddisfly. The case is usually portable, dragged around like a snail shell as the insect moves, and held in place by a pair of hooked prolegs at the tip of the abdomen. Most species have thread-like abdominal gills and get oxygen from water that circulates inside the case. All larval growth and development (including pupation) occurs within the case.
Lepidoptera
 Lepidoptera (moths and butterflies) is the second largest order in the class Insecta. Adults are distinctive for their large wings (relative to body size) which are covered with minute overlapping scales. Most entomologists believe that these scales are structurally related to the hair (setae) covering adult caddisflies. Lepidopteran wing scales often produce distinctive color patterns that play an important role in courtship and intraspecific recognition. In flight, front and hind wings are linked together by a bristle (frenulum) or a membranous flap (jugum) so both wings move up and down in synchrony.
Lepidoptera (moths and butterflies) is the second largest order in the class Insecta. Adults are distinctive for their large wings (relative to body size) which are covered with minute overlapping scales. Most entomologists believe that these scales are structurally related to the hair (setae) covering adult caddisflies. Lepidopteran wing scales often produce distinctive color patterns that play an important role in courtship and intraspecific recognition. In flight, front and hind wings are linked together by a bristle (frenulum) or a membranous flap (jugum) so both wings move up and down in synchrony.
Although moths probably diverged from caddisflies in the early Triassic period, about 230 million years ago, adults in a few primitive families (e.g.. Micropterygidae) still retain evidence of chewing mouthparts. In all other lepidopteran families, the mouthparts are vestigal or form a tubular proboscis that lies coiled like a watch spring beneath the head. This proboscis is derived from portions of the maxillae. It uncoils by hydrostatic pressure and acts as a siphon tube for sipping liquid nutrients, such as nectar, from flowers and other substrates.
From a taxonomic standpoint, the distinction between moths and butterflies is largely artificial — some moths are more similar to butterflies than to other moths. As a rule, butterflies are diurnal, brightly colored, and have knobs or hooks at the tip of the antennae. At rest, the wings are held vertically over the body. In contrast, most (but not all) moths are nocturnal. They are typically drab in appearance, and have thread-like, spindle-like, or comb-like antennae. At rest, their wings are held horizontally against the substrate, folded flat over the back, or curled around the body.
Nearly all lepidopteran larvae are called caterpillars. More about
Lepidoptera They have a well-developed head with chewing mouthparts. In addition to three pairs of legs on the thorax, they have two to eight pairs of fleshy abdominal prolegs that are structurally different from the thoracic legs. Most lepidopteran larvae are herbivores; some species eat foliage, some burrow into stems or roots, and some are leaf-miners.
Hymenoptera
 The Hymenoptera (sawflies, ants, wasps, and bees) is a distinctive group of insects that has relatively few advanced characteristics in common with other endopterygote groups. Although the fossil record for Hymenoptera dates back only to the Triassic period (220-230 million years ago), we suspect that these insects first appeared in the Permian period (225-290 million years ago) as an early offshoot of the mecopteran lineage. This interpretation is supported by the fact that sawflies and horntails, the most primitive members of the order, produce silk from modified salivary glands, have abdominal prolegs in the larval stage, and show reduction in size of the hind wings. More advanced groups (wasps, bees, and ants) appeared much later in the fossil record (mid-Jurassic) and probably co-evolved with flowering plants during the Cretaceous period.
The Hymenoptera (sawflies, ants, wasps, and bees) is a distinctive group of insects that has relatively few advanced characteristics in common with other endopterygote groups. Although the fossil record for Hymenoptera dates back only to the Triassic period (220-230 million years ago), we suspect that these insects first appeared in the Permian period (225-290 million years ago) as an early offshoot of the mecopteran lineage. This interpretation is supported by the fact that sawflies and horntails, the most primitive members of the order, produce silk from modified salivary glands, have abdominal prolegs in the larval stage, and show reduction in size of the hind wings. More advanced groups (wasps, bees, and ants) appeared much later in the fossil record (mid-Jurassic) and probably co-evolved with flowering plants during the Cretaceous period.
As a rule, members of the order Hymenoptera can be regarded as ecological specialists. Most species are rather narrowly adapted to specific habitats and/or specific hosts. Their remarkable success as a taxon probably has more to do with their immense range of behavioral adaptation rather than any physical or biochemical characteristic. The Hymenoptera is the only order besides the Isoptera (termites) to have evolved complex social systems with division of labor.
Herbivory is common among the primitive Hymenoptera (suborder Symphyta), in the gall wasps (Cynipidae), and in some of the ants and bees. Most other Hymenoptera are predatory or parasitic. The large hunting wasps are agile predators that catch and paralyze insects (or spiders) as food for their offspring. The greatest diversity, though, is found among the many families of parasitoid wasps whose larvae feed internally on the living tissues of other arthropods (or their eggs). These insects eventually kill their host, but not before completing their own larval development within its body. Despite their small size and characteristically narrow host range, these wasps are highly abundant and exert a tremendous impact on the population dynamics of many other insect species.
Most of the Hymenoptera have relatively unspecialized mandibulate mouthparts. More about
Hymenoptera An exception is found in the bees (superfamily Apidoidea) where the maxillae and labium are modified into a proboscis that works like a tongue to collect nectar from flowers. In these insects, the mandibles are used to gather or manipulate pollen and wax.
Except for worker ants, most adult Hymenoptera have two pairs of wings. Front and hind wings are linked together by hooks (hamuli) along the leading edge of the hind wings that catch in a fold near the back of the front wings. In flight, both wings operate in unison to form a single aerodynamic surface.

{kind=link}
{kind=link}