
The Paraneopteran Orders
Although it contains fewer orders, the Paraneoptera branch of the insect’s phylogenetic tree encompasses nearly three times as many species as the Polyneoptera branch and has far greater ecological and economic impact. All of the insects classified here exhibit various “reductions” or “simplifications” from the primitive body-plan found in typical polyneopterans . Cerci, for example, are entirely absent in all living paraneopterans (some taxonomists classify this group as the superorder Acercaria, meaning without cerci). Other “reductions” occur in wing venation, in the number of tarsal segments (not more than three), in the number of Malpighian tubules (not more than six), and in the number of ganglia present in the ventral nerve cord (not more than five).
Perhaps the most significant hallmark of the paraneopteran lineage is a progressive development of haustellate mouthparts, variously adapted for scraping food from a substrate or gathering it by suction in liquid or semi-liquid form. Mouthparts of this type are distinctive because one or more of their structural components is elongated to form a rod or a stylet. In their most specialized form, as in the order Hemiptera, haustellate mouthparts form a tubular rostrum (also called a beak or a proboscis) with feeding stylets that can pierce the tissue of a plant or animal host and suck out the liquid contents. Enlargement of cibarial muscles (along the anterior surface of the pharynx) usually accompanies the development of haustellate mouthparts. Contraction of these muscles creates the bellows-like suction that draws food up through the proboscis (see illustration on the apterygota page ).
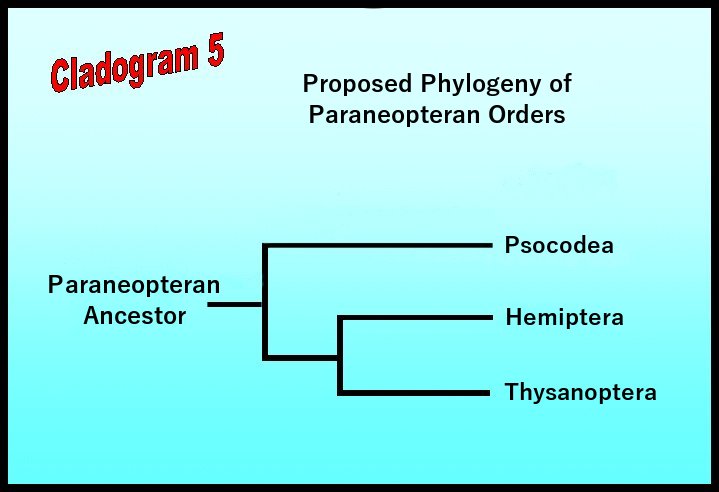
Entomologists generally agree that members of the Paraneoptera complex arose from a common ancestor during the Carboniferous period, around 290 million years ago. Adaptive radiation within this lineage eventually led to the evolution of two sister groups (Cladogram 5): one giving rise to the present-day booklice, barklice, and parasitic lice (Psocodea); the other giving rise to thrips, true bugs, aphids, leafhoppers, etc. (Thysanoptera and Hemiptera).
Psocodea
 The order Psocodea contains the booklice, barklice, and parasitic lice. In the past, members of this order have been grouped in several ways and classified in various orders, including Corrodentia (barklice), Psocoptera (barklice and booklice), Phthiraptera (parasitic lice), Mallophaga (biting or chewing lice), and Anoplura (sucking lice).
The order Psocodea contains the booklice, barklice, and parasitic lice. In the past, members of this order have been grouped in several ways and classified in various orders, including Corrodentia (barklice), Psocoptera (barklice and booklice), Phthiraptera (parasitic lice), Mallophaga (biting or chewing lice), and Anoplura (sucking lice).
Booklice and barklice are often regarded as the most primitive paraneopterans alive today because their mouthparts show the least modification from a primitive mandibulate condition. In fact, only the lacinia (a subdivision of the maxilla) has become a separate, rod-like structure that is pushed against the substrate as a brace while the mandibles scrape off surrounding food particles. The pharynx and hypopharynx are also modified for grinding food in a mortar-and-pestle arrangement.
Barklice generally live in moist terrestrial environments (in leaf litter, beneath stones, on vegetation, or under the bark of trees) where they forage on algae, lichens, fungi, and various plant products. They may grow to 10 mm in length and are frequently winged during the adult stage. Some species are gregarious. They often live in small colonies beneath a gossamer web spun with silk from their labial glands. Booklice are more common in human dwellings and warehouses. They are wingless and much smaller than barklice (less than 2 mm). Most species feed on stored grains, book bindings, wallpaper paste, fabric sizing, and other starchy products.
Although most barklice are free-living, a few genera live in the nests of birds. More about
Psocodea They survive by feeding on residues of feathers or skin cells, but never on the birds themselves. Most entomologists suspect that parasitic lice (ectoparasities of birds and mammals) evolved directly from these commensal barklice. Even today, it is possible to find a gradual progression of species with increasing dependence on vertebrate hosts. A close phylogenetic relationship between barklice and parasitic lice is also supported by similarities in the structure of mouthparts (particularly the hypopharynx).
 Parasitic lice can be further subdivided into biting lice and sucking lice. The distinction is based primarily on the presence or absence of mandibles that are suitable for biting and chewing. All of these parasitic lice are external parasites of birds and mammals. The biting lice probably evolved first on birds, feeding on feathers and dead skin cells. But sometime during the Cretaceous period (less than 135 million years ago) biting lice expanded their host range to include certain groups of mammals. A few of these lice developed the habit of breaking their host’s skin and feeding on its blood. This lineage presumably gave rise to the sucking lice, all of which are blood-feeding ectoparasites of placental mammals.
Parasitic lice can be further subdivided into biting lice and sucking lice. The distinction is based primarily on the presence or absence of mandibles that are suitable for biting and chewing. All of these parasitic lice are external parasites of birds and mammals. The biting lice probably evolved first on birds, feeding on feathers and dead skin cells. But sometime during the Cretaceous period (less than 135 million years ago) biting lice expanded their host range to include certain groups of mammals. A few of these lice developed the habit of breaking their host’s skin and feeding on its blood. This lineage presumably gave rise to the sucking lice, all of which are blood-feeding ectoparasites of placental mammals.
Unlike many other ectoparasites, sucking lice cannot survive long if separated from the body of their host. Eggs (called nits) are glued directly to the hair or feathers and nymphs feed on the parental host. Since lice have no wings, dispersal to new host animals is limited to occasions when members of the host species come into direct contact with each other. This close interspecific association means that most lice are limited to a very narrow host range — often only a single species.
Thysanoptera
 A second branch of the Paraneoptera lineage includes insects in which the haustellate mouthparts contain feeding stylets derived from both the mandibles and maxillae. In Thysanoptera (thrips), there are three needle-like stylets that form an asymmetrical feeding apparatus housed within a conical mouth opening. The stylets (two maxillae and one mandible) pierce and lacerate host tissue. Macerated food is then drawn into the mouth by suction from the cibarial pump.
A second branch of the Paraneoptera lineage includes insects in which the haustellate mouthparts contain feeding stylets derived from both the mandibles and maxillae. In Thysanoptera (thrips), there are three needle-like stylets that form an asymmetrical feeding apparatus housed within a conical mouth opening. The stylets (two maxillae and one mandible) pierce and lacerate host tissue. Macerated food is then drawn into the mouth by suction from the cibarial pump.
Thrips are generally small insects (under 3 mm). Most species feed on plant tissues (often in flower heads), but some are predators of mites and various small insects (including other thrips). Many species are parthenogenetic. Adults may be winged or wingless. When present, the wings are slender and rod-like with a dense fringe of long hairs.
Although Thysanoptera are hemimetabolous, More about
Thysanoptera many species undergo an extended metamorphosis in which the final immature stage is quiescent, non-feeding, and sometimes even enclosed in a silken cocoon. This developmental stage, usually called a “pupa”, has aroused a great deal of speculation by some entomologists who claim that thrips represent an “intermediate” stage between hemi- and holometabolous development. A close examination of the thysanopteran “pupa”, however, reveals that it does not undergo any internal transformation. Without additional evidence to support a phylogenetic link to the Holometabola, it would appear that this “pupal stage” may be nothing more than a curious coincidence of convergent evolution.
Hemiptera
 Hemiptera is the final order in the Paraneoptera complex and by far the largest, both in number of species and in ecological importance. All of the insects classified as Hemiptera have a highly specialized rostrum (proboscis) in which all structural elements are elongated to form a tubular feeding channel. The mandibles and maxillae are long and thread-like. All four of these feeding stylets interlock to form a flexible feeding tube that is no more than 0.1 mm in diameter yet contains both a food channel and a salivary channel. The stylets are enclosed within a protective sheath (the labium) that shortens or retracts during feeding.
Hemiptera is the final order in the Paraneoptera complex and by far the largest, both in number of species and in ecological importance. All of the insects classified as Hemiptera have a highly specialized rostrum (proboscis) in which all structural elements are elongated to form a tubular feeding channel. The mandibles and maxillae are long and thread-like. All four of these feeding stylets interlock to form a flexible feeding tube that is no more than 0.1 mm in diameter yet contains both a food channel and a salivary channel. The stylets are enclosed within a protective sheath (the labium) that shortens or retracts during feeding.
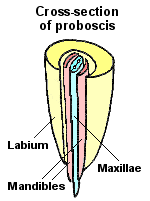 Not long after these piercing/sucking mouthparts evolved, the hemipteran lineage seems to have split into two sister groups. In one group, Homoptera, the rostrum is relatively short (1-3 segments) and emerges from near the ventral posterior margin of the head. These insects (e.g., leafhoppers, cicadas, aphids, etc.) are said to have opistognathous mouthparts (from the Greek “opisto-” meaning backward and “gnatha” meaning mouth). Although some Homoptera are secondarily wingless, the majority have membranous or uniformly textured wings that fold tent-like over the body at rest.
Not long after these piercing/sucking mouthparts evolved, the hemipteran lineage seems to have split into two sister groups. In one group, Homoptera, the rostrum is relatively short (1-3 segments) and emerges from near the ventral posterior margin of the head. These insects (e.g., leafhoppers, cicadas, aphids, etc.) are said to have opistognathous mouthparts (from the Greek “opisto-” meaning backward and “gnatha” meaning mouth). Although some Homoptera are secondarily wingless, the majority have membranous or uniformly textured wings that fold tent-like over the body at rest.
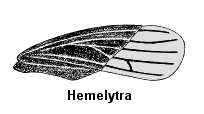 In members of the second group, Heteroptera, the rostrum is relatively long (3-4 segments) and arises near the front or lower front of the head (prognathous or hypognathous). These insects are known as the “true bugs”. They have very distinctive front wings, called hemelytra, in which the basal half is leathery and the apical half is membranous. At rest, these wings cross over one another to lie flat along the insect’s back.
In members of the second group, Heteroptera, the rostrum is relatively long (3-4 segments) and arises near the front or lower front of the head (prognathous or hypognathous). These insects are known as the “true bugs”. They have very distinctive front wings, called hemelytra, in which the basal half is leathery and the apical half is membranous. At rest, these wings cross over one another to lie flat along the insect’s back.
For many years, the ordinal status of Heteroptera and Homoptera has been a source of great controversy among entomologists. Although “splitters” still prefer to divide them into two separate orders, a majority of taxonomists now classify Heteroptera and Homoptera as suborders within Hemiptera. Since the order is large and diverse, we will often use suborder names when referring to characteristics that are not common to both groups.
The Heteroptera is an unusually diverse assemblage of insects. More about
Heteroptera Members of this suborder have become adapted to a broad range of habitats — terrestrial, aquatic, and semi-aquatic. Terrestrial species are often associated with plants. They feed in vascular tissues or on the nutrients stored within seeds. Other species live as scavengers in the soil or underground in caves or ant nests. Still others are predators on a variety of small arthropods.
A few species even feed on the blood of vertebrates. Bed bugs, and other members of the family Cimicidae, live exclusively as ectoparasites on birds and mammals (including humans). Aquatic Heteroptera can be found on the surface of both fresh and salt water, near shorelines, or beneath the water surface in nearly all freshwater habitats. With only a few exceptions, these insects are predators of other aquatic organisms.
 All members of the suborder Homoptera feed by withdrawing sap from vascular plants. But beyond this, it is difficult to generalize about the biology of these insects. Cicadas are the largest members of the suborder. As nymphs, they live underground and feed on the roots of trees and shrubs. Some species complete development in as little as four years, but others have a 13- or 17-year life cycle. In contrast, the aphids are tiny, soft-bodied insects with multiple generations per year. Many species have complex life cycles involving more than one host plant. Winged and wingless forms of the same species may develop at different times of the year. Asexual reproduction (parthenogenesis) is common and males are unknown in some species. The scale insects are even more specialized. During much of their life cycle, they remain immobile, living beneath an impervious cover of wax or cuticle that they secrete over themselves. Legs and antennae often disappear after the first molt. Only newly hatched nymphs and adult males bear any resemblance to other insects. Females grow to sexual maturity, mate, produce off-spring, and die without ever leaving their protective cover.
All members of the suborder Homoptera feed by withdrawing sap from vascular plants. But beyond this, it is difficult to generalize about the biology of these insects. Cicadas are the largest members of the suborder. As nymphs, they live underground and feed on the roots of trees and shrubs. Some species complete development in as little as four years, but others have a 13- or 17-year life cycle. In contrast, the aphids are tiny, soft-bodied insects with multiple generations per year. Many species have complex life cycles involving more than one host plant. Winged and wingless forms of the same species may develop at different times of the year. Asexual reproduction (parthenogenesis) is common and males are unknown in some species. The scale insects are even more specialized. During much of their life cycle, they remain immobile, living beneath an impervious cover of wax or cuticle that they secrete over themselves. Legs and antennae often disappear after the first molt. Only newly hatched nymphs and adult males bear any resemblance to other insects. Females grow to sexual maturity, mate, produce off-spring, and die without ever leaving their protective cover.
In most of the Homoptera, a portion of the digestive system is modified into a filter chamber. More about
Homoptera This structure allows the insects to ingest and process large volumes of plant sap. Excess water, sugars, and certain amino acids bypass most of the midgut and are shunted directly into the hindgut for excretion as honeydew. Only a small volume of filtered plant sap passes through the midgut for digestion and absorption. Many species of ants are attracted by the honydew and provide provide care and protection for the homopterans in exchange for the honeydew they excrete.

{kind=link}