
An insect’s exoskeleton (integument) serves not only as a protective covering over the body, but also as a surface for muscle attachment, a water-tight barrier against desiccation, and a sensory interface with the environment. It is a multi-layered structure with four functional regions: epicuticle, procuticle, epidermis, and basement membrane.
The epidermis is primarily a secretory tissue formed by a single layer of epithelial cells. It is responsible for producing at least part of the basement membrane as well as all of the overlying layers of cuticle. The basement membrane is a supportive bilayer of amorphous mucopolysaccharides (basal lamina) and collagen fibers (reticular layer). The membrane serves as a backing for the epidermal cells and effectively separates the hemocoel (insect’s main body cavity) from the integument.
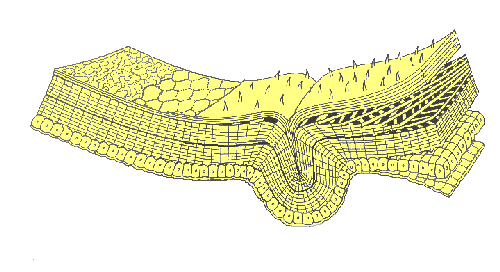 The procuticle lies immediately above the epidermis. It contains microfibers of chitin surrounded by a matrix of protein that varies in composition from insect to insect and even from place to place within the body of a single insect.
The procuticle lies immediately above the epidermis. It contains microfibers of chitin surrounded by a matrix of protein that varies in composition from insect to insect and even from place to place within the body of a single insect.
As the procuticle forms, it is laid down in thin lamellae with chitin microfibers oriented at a slightly different angle in each subsequent layer. In some parts of the body, procuticle stratifies into a hard, outer exocuticle and a soft, inner endocuticle.
Differentiation of exocuticle involves a chemical process (called sclerotization) that occurs shortly after each molt. During sclerotization, individual protein molecules are linked together by quinone compounds. These reactions “solidify” the protein matrix, creating rigid “plates” of exoskeleton known as sclerites. Quinone cross-linkages do not form in parts of the exoskeleton where resilin (an elastic protein) is present in high concentrations. These areas are membranes — they remain soft and flexible because they never develop a well-differentiated exocuticle.
The epicuticle is the outermost part of the cuticle. Its function is to reduce water loss and block the invasion of foreign matter. The innermost layer of epicuticle is often called the cuticulin layer, a stratum composed of lipoproteins and chains of fatty acids embedded in a protein-polyphenol complex. An oriented monolayer of wax molecules lies just above the cuticulin layer; it serves as the chief barrier to movement of water into or out of the insect’s body. In many insects a cement layer covers the wax and protects it from abrasion.
In many insects, certain epidermal cells are specialized as exocrine glands. These large, secretory cells produce compounds (e.g. pheromones, repellants, etc.) that are released on the surface of the exoskeleton through microscopic ducts.
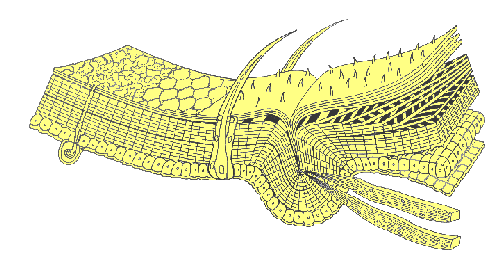 Tiny hair-like projections or surface sculpturing of the cuticle are known as microtrichae or pile (PILL-EE). These acellular structures consist of a solid core of exocuticle covered by a thin layer of epicuticle. Larger hairs, bristles, and scales (called setae or macrotrichae) are the product of two specialized epidermal cells: a trichogen cell (the hair shaft) and a tormogen cell (the socket). Multicellular projections of the exoskeleton are called spines (or spurs, if movable). They are lined with epidermis and contain both procuticle and epicuticle.
Tiny hair-like projections or surface sculpturing of the cuticle are known as microtrichae or pile (PILL-EE). These acellular structures consist of a solid core of exocuticle covered by a thin layer of epicuticle. Larger hairs, bristles, and scales (called setae or macrotrichae) are the product of two specialized epidermal cells: a trichogen cell (the hair shaft) and a tormogen cell (the socket). Multicellular projections of the exoskeleton are called spines (or spurs, if movable). They are lined with epidermis and contain both procuticle and epicuticle.
Skeletal muscles attach to the inner surface of the integument. Despite small body size, insects have many more muscles than vertebrates because the exoskeleton affords a larger surface area than an endoskeleton (relative to body volume) for muscle attachment. An insect owes its incredible strength to the geometry of its musculature — providing optimal leverage for movement of appendages.
Invaginations (inward folds) of the exoskeleton add to its strength and rigidity. They also provide increased surface area for attachment of muscles. Ridge-like invaginations are called apodemes. They are usually visible externally as a groove (suture). Finger-like invaginations are called apophyses. A tiny pit usually marks their location externally.
The colors found in the integument of insects are produced either by pigment molecules, usually located in the cuticle, or by physical characteristics of the integument that cause scattering, interference, or diffraction of light. Pigments that are frequently present include the pterines, melanins, carotenoids, and mesobiliverdin.
Color patterns may change over time. Rapid, temporary changes may occur in response to daily environmental conditions or to the threat of danger. Slower, more permanent changes are usually related to seasonal changes in the environment or hormonal influences.
