Embryogenesis
Embryogenesis is a developmental process that usually begins once the egg has been fertilized. It involves multiplication of cells (by mitosis) and their subsequent growth, movement, and differentiation into all the tissues and organs of a living insect. The field of insect embryology has recently yielded stunning insights into the developmental processes of humans and other vertebrate organisms. There is remarkable similarity in genes responsible for organizing the fundamental body plan in vertebrates and invertebrates. For example, eyeless, a gene needed for development of an insect’s compound eyes is also necessary for development of a mouse’s vertebrate eyes!
Although much of insect embryology is still a mystery, there has been remarkable progress in knowledge over the past few years thanks to new methods in molecular biology and genetic engineering. Fruit flies, silkworms, and hornworms are proving to be a “rosetta stone” for embryology. An insect’s egg is much too large and full of yolk to simply divide in half like a human egg during its initial stages of development (imagine how much time and energy it would take just to build new cell membranes!). Birds have this same problem — think of the yolk in a chicken’s egg. Birds solve the problem by having the embryo develop within a tiny spot of cytoplasm (the blastodisc) on the surface of the yolk. Insects solve the problem by “cloning” the zygote nucleus (mitosis without cytokinesis) through 12-13 division cycles to yield about 5000 daughter nuclei. This process of nuclear division is known as superficial cleavage (in “true” cleavage entire cells divide). As they form, the cleavage nuclei (often called “energids”) migrate through the yolk toward the perimeter of the egg. They settle in the band of periplasm where they engineer the construction of membranes to form individual cells. The end result of “cleavage” is the blastoderm — a one-cell-thick layer of cells surrounding the yolk.
The first cleavage nuclei to reach the vicinity of the öosome are “reserved” for future reproductive purposes — they do not travel to the periplasm and do not form any part of the blastoderm. Instead, they stop dividing and form germ cells that remain segregated throughout much of embryogenesis. These cells will eventually migrate into the developing gonads (ovaries or testes) to become primary öocytes or spermatocytes. Only when the adult insect finally reaches sexual maturity will these cells begin dividing (by meiosis) to form gametes of the next generation (eggs or sperm). Germ cells never grow or divide during embryogenesis, so DNA for the next generation is “conserved” from the very beginning of development. This strategy has a clear selective advantage: it minimizes the risk that an error in replication (a genetic defect) will accidently be passed on to the next generation.
Blastoderm cells on one side of the egg begin to enlarge and multiply. This region, known as the germ band (or ventral plate), is where the embryo’s body will develop. The rest of the cells in the blastoderm become part of a membrane (the serosa) that forms the yolk sac. Cells from the serosa grow around the germ band, enclosing the embryo in an amniotic membrane.
At this stage of development, when the embryo is not much more than a single layer of cells, a group of control genes (called homeotic selector genes) become active. These genes encode for proteins that contain a special active site (the homeobox) for binding with DNA. They interact with specific locations in the genome where they function as switches for activating (or inhibiting) the expression of other genes. Basically, each selector gene controls the expression of certain other genes within a restricted domain of cells based on their location in the germ band.
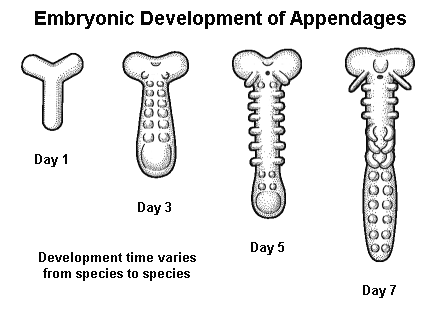
By regulating activity within a suite of genes that produce hormone-like “organizer” chemicals, cell-surface receptors, and structural elements, the selector genes guide the development of individual cells and channel them into different “career paths”. This process, called differentiation, continues until the fundamental body plan is mapped out — first into general regions along an anterio-posterior axis, then into individual segments, and finally into specialized structures or appendages.