
Insect Defenses
For many insects, a quick escape by running or flying is the primary mode of defense. A cockroach, for example, has mechanoreceptive hairs (setae) on the cerci that are sensitive enough to detect the change in air pressure that precedes a fast moving object (like your foot). Nerve impulses from these receptors travel through giant neurons to thoracic ganglia at speeds up to 3 meters per second, triggering an evasive response by the legs in less than 50 milliseconds. House flies have a similar reaction time when you try to swat them. They leap into the air and begin flapping their wings 30-50 milliseconds after sensing a threat.
Tiger moths (family Arctiidae) can detect ultrasonic echolocation by bats. At low intensity, they fly away from the bat, but if the bat’s call increases to a certain threshold they quickly drop from the air in an evasive, looping dive. Other alarm reactions may be less dramatic, but just as effective: Madagascar cockroaches hiss when disturbed; cuckoo wasps curl up into hard, rigid balls; tortoise beetles have strong adhesive pads on their tarsi and hold themselves tight and flat against a leaf or stem. Other insects simply “play dead” (thanatosis) — they release their grip on the substrate and fall to the ground where they are hard to find as long as they remain motionless.
An insect’s hard exoskeleton may serve as an effective defense against some predators and parasites. Large weevils are notorious for their hard bodies – as you may discover for yourself the first time you bend an insect pin trying to push it through the thorax. Most diving beetles are hard, slick, and streamlined; even if you can catch them, they will often squirm out of your grip.
 Spines, bristles, and hairs may be effective mechanical deterrents against predators and parasites. A mouthful of hair can be an unpleasant experience for a predator and parasitic flies or wasps may have a hard time getting close enough to the insect’s body to lay their eggs. Some caterpillars incorporate body hairs into the silk of their cocoon as an additional defense against predation.
Spines, bristles, and hairs may be effective mechanical deterrents against predators and parasites. A mouthful of hair can be an unpleasant experience for a predator and parasitic flies or wasps may have a hard time getting close enough to the insect’s body to lay their eggs. Some caterpillars incorporate body hairs into the silk of their cocoon as an additional defense against predation.
Some insects have a “fracture line” in each appendage (often between the trochanter and the femur) that allows a leg to break off easily if it is caught in the grasp of a predator. This phenomenon, called autotomy, is most common in crane flies, walkingsticks, grasshoppers, and other long-legged insects. In most cases, sacrificing a limb in this manner creates only a minor disability. In fact, walkingsticks (especially young nymphs) may regenerate all or part of a missing appendage over the course of several molts.
Chemical Defenses
Many insects are equipped to wage chemical warfare against their enemies. In some cases, they manufacture their own toxic or distasteful compounds. In other cases, the chemicals are acquired from host plants and sequestered in the hemolymph or body tissues. When threatened or disturbed, the noxious compounds may be released onto the surface of the body as a glandular ooze, into the air as a repellent volatile, or aimed as a spray directly at the offending target.
Defensive chemicals typically work in one of four ways:
-
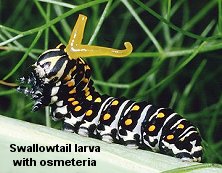
- Repellency — a foul smell or a bad taste is often enough to discourage a potential predator. Stink bugs, for example, have specialized exocrine glands located in the thorax or abdomen that produce foul-smelling hydrocarbons. These chemicals accumulate in a small reservoir adjacent to the gland and are released onto the body surface only as needed. The larvae of certain swallowtail butterflies have eversible glands, called osmeteria, located just behind the head. When a caterpillar is disturbed, it rears up, everts the osmeteria to release a repellent volatile, and waves its body back and forth to ward off intruders.
- Induce cleaning — irritant compounds often induce cleaning behavior by a predator, giving the prey time to escape. Some blister beetles (family Meloidae) produce cantharidin, a strong irritant and blistering agent that circulates in their hemolymph. Droplets of this blood ooze from the beetle’s leg joints when it is disturbed or threatened — an adaptation known as reflex bleeding. Irritant sprays are produced by some termites, cockroaches, earwigs, stick insects, and beetles. The notorious bombardier beetles store chemical precursors for an explosive reaction mixture in specialized glands. When threatened, these precursors are mixed together to produce a forceful discharge of boiling hot benzoquinone and water vapor (steam).
- Adhesion — sticky compounds that harden like glue to incapacitate an attacker. Several species of cockroach guard their backsides with a slimy anal secretion that quickly cripples any worker ants that launch an attack. Similarly, members of the soldier caste in nasute termites have nozzle-like heads equipped with a defensive gland that can shoot a cocktail of defensive chemicals at intruders. The compounds, which are both irritating and immobilizing, have been shown to be highly effective against ants, spiders, centipedes, and other predatory arthropods.
 Cause pain or discomfort — Saddleback caterpillars, larvae of the io moth, and various other Lepidopteran larvae have hollow body hairs that contain a painful irritant. Simply brushing against these urticating hairs will cause them to break and release their contents onto your skin. The consequence is an intense burning sensation that may last for several hours. Many ants, bees, and wasps (the aculeate Hymenoptera) deliver venom to their enemies by means of a formidable stinger (modified ovipositor). The venom is a complex mixture of proteins and amino acids that not only induces intense pain but may also trigger an allergic reaction in the victim.
Cause pain or discomfort — Saddleback caterpillars, larvae of the io moth, and various other Lepidopteran larvae have hollow body hairs that contain a painful irritant. Simply brushing against these urticating hairs will cause them to break and release their contents onto your skin. The consequence is an intense burning sensation that may last for several hours. Many ants, bees, and wasps (the aculeate Hymenoptera) deliver venom to their enemies by means of a formidable stinger (modified ovipositor). The venom is a complex mixture of proteins and amino acids that not only induces intense pain but may also trigger an allergic reaction in the victim.
Protective Coloration
Biologists recognize that there is usually an underlying rationale for the great diversity of shapes and colors found in the insect world. We may not know why a particular species has parallel ridges on the pronotum or black spots on the wings, but we can be reasonably certain that this shape or color has contributed in some way, however small, to the overall fitness of the species. It is obvious that at least some of the colors and patterns serve a defensive function by offering a degree of protection from predators and parasites. These patterns, collectively known as protective coloration, fall into four broad categories:
-
- Crypsis — Insects that blend in with their surroundings often manage to escape detection by predators and parasites. This tactic, called cryptic coloration, involves not only matching the colors of the background but also disrupting the outline of the body, eliminating reflective highlights from smooth body surfaces, and avoiding sudden movements that might betray location. Obviously, this tactic loses much of its effectiveness if an insect moves from one type of habitat to another. Well-camouflaged insects usually stay close to home or make only short trips and return quickly to the shelter of their protective cover. Many ground-dwelling grasshoppers and katydids, for example, have colors of mottled gray and brown that help them “disappear” against a background of dried leaves or gravel. On the other hand, closely related species that live in foliage are usually a shade of green that matches the surrounding leaves. The larvae of some lacewings improve their camouflage by attaching bits of moss or lichen from their environment onto the dorsal side of their body. Examples of Crypsis
- Mimesis — Some insects “hide in plain sight” by resembling other objects in the environment. A thorn could really be a treehopper; a small twig might be a walkingstick, an assassin bug, or the caterpillar of a geometrid moth; and sometimes a dead leaf turns out to be a katydid, a moth, or even a butterfly. This “mimicry” of natural objects is often known as mimesis. It goes far beyond imitation of plant parts:
- Some swallowtail larvae resemble bird droppings, others have false eyespots on the thorax that create a convincing imitation of a snake’s head.
- The likeness of a caterpillar can be found on the outer edge of many lepidopteran wings, perhaps serving to fool predatory birds that may peck at the wing margin instead of the butterfly’s body.
- Many butterflies and moths have eyespots on the wings that emulate the face of an owl or some other large animal.
- Slug caterpillars and hag moth larvae look like hair balls or small furry mammals.
- Warning Colors — Insects that have an active means of defense (like a sting or a repellent spray) frequently display bright colors or contrasting patterns that tend to attract attention. These visually conspicuous insects illustrate aposematic coloration, a term derived from the Greek words apo- (from a distance) and sema (a sign or signal) — meaning “a signal from afar”. A predator quickly learns to associate the distinctive coloration with an “unpleasant” outcome, and one such encounter is usually enough to insure avoidance of that prey in the future. A few individuals will die as sacrifices, but for the species as a whole, it pays to advertise!
- Mimicry — If a distinctive visual appearance is sufficient to protect an unpalatable insect from predation, then it stands to reason that other insects might also avoid predation by adopting a similar appearance. This ploy, essentially a form of “false advertising”, was first recognized and described by Henry W. Bates in 1861. Today, it is commonly known as Batesian mimicry. Viceroy butterflies (mostly palatable to birds) are largely protected from predation because they resemble monarch butterflies (very distasteful). Many species of bee flies, flower flies, robber flies, and clear-winged moths are similarly protected because they mimic the appearance (and often the behavior) of stinging bees and wasps. Batesian mimicry is usually a successful strategy as long as the model and mimic are found in the same location, the mimic’s population size is smaller than that of the model, and predators associate the model’s appearance with an unpleasant effect.In 1879, Fritz Müller recognized that two or more distasteful species often share the same aposematic color patterns. Many species of wasps, for example, have alternating bands of black and yellow on the abdomen. This defensive tactic, commonly known as Müllerian mimicry, benefits all members of the group because it spreads the liability for “educating the predator” over more than one species. In fact, as the number of species in a Müllerian complex increases, there is a greater selective advantage for each individual species.
 Mimicry has been carried to extremes in some tropical Lepidoptera where both related and unrelated species resemble each other in size, shape, color, and wing pattern. Collectively, these butterflies (and sometimes moths) form mimicry rings that may include both palatable and unpalatable species. In South America, for example, longwing butterflies (Family Nymphalidae) form a mimicry ring that includes at least twelve different species (including one moth).
Mimicry has been carried to extremes in some tropical Lepidoptera where both related and unrelated species resemble each other in size, shape, color, and wing pattern. Collectively, these butterflies (and sometimes moths) form mimicry rings that may include both palatable and unpalatable species. In South America, for example, longwing butterflies (Family Nymphalidae) form a mimicry ring that includes at least twelve different species (including one moth).
Although natural selection favors individuals in a population with the best camouflage or mimicry, it also favors the predator or parasite with the best prey-finding acumen. As a result of these competing interests, coevolution between predator and prey populations inevitably leads to an ongoing escalation of offensive and defensive measures — a scenario that Leigh Van Valen of Chicago University describes as an evolutionary “arms race”.
 In order to survive in the arms race, both predator and prey must constantly evolve in response to the other’s changes. Failure to “keep up” concedes a competitive advantage to the opponent and may lead to extinction.
In order to survive in the arms race, both predator and prey must constantly evolve in response to the other’s changes. Failure to “keep up” concedes a competitive advantage to the opponent and may lead to extinction.
The idea that perpetual change is necessary just to maintain the status quo has been coined the Red Queen’s Hypothesis. This name refers to a scene from the stories of Alice in Wonderland by Lewis Carroll. In Through the Looking Glass, Alice meets a chess piece, the Red Queen. After running hard to follow the Queen, Alice discovers that she has not moved from where she started. Asked about this paradox, the Red Queen replies, “Here, you see, it takes all the running you can do to keep in the same place.”
