
The Arthropods
The First Arthropods
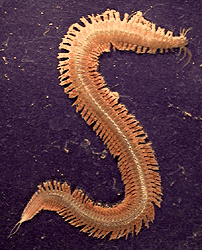 A variety of marine worms (Annelida and Protoannelida) lived in the ocean sediments during the Cambrian period. These creatures were bilaterally symmetrical, soft-bodied, and multisegmented. They had no distinct head capsule and lacked both eyes and antennae. Some species may have had lobe-like lateral appendages similar to the parapodia of polychaete worms that thrive in the muddy sediments of today’s ocean floor. But structural differences between the lateral appendages of polychaetes and early arthropods suggest that these two groups diverged from a common annelid ancestor some 500-600 million years ago. All annelids were soft-bodied. They decayed rapidly after death and were not well-preserved in the fossil record.
A variety of marine worms (Annelida and Protoannelida) lived in the ocean sediments during the Cambrian period. These creatures were bilaterally symmetrical, soft-bodied, and multisegmented. They had no distinct head capsule and lacked both eyes and antennae. Some species may have had lobe-like lateral appendages similar to the parapodia of polychaete worms that thrive in the muddy sediments of today’s ocean floor. But structural differences between the lateral appendages of polychaetes and early arthropods suggest that these two groups diverged from a common annelid ancestor some 500-600 million years ago. All annelids were soft-bodied. They decayed rapidly after death and were not well-preserved in the fossil record.
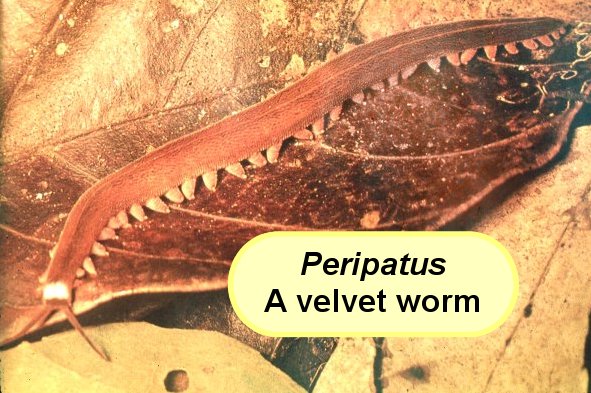 At one time, the Onychophora or velvet worms were suggested as a possible evolutionary link (intermediate) between the annelids and the arthropods. These mostly-tropical invertebrates, with only 75 extant species, live on the forest floor among moist, decaying leaves and feed on a mixed diet of plant and animal tissues. Like annelid worms, the Onychophorans have segmented bodies containing paired excretory organs (nephridia) and a combination of both male and female sex organs (monoecious). They lack a true exoskeleton, but the body is covered by a soft, chitinous cuticle. In true arthropod fashion, this cuticle is periodically shed (molted) to allow for continued growth. Onychophorans also share numerous other characteristics with the arthropods, including antennae, an open circulatory system, walking legs with claws, paired mandibles, and a system of slender air tubes (tracheae) for respiration.
At one time, the Onychophora or velvet worms were suggested as a possible evolutionary link (intermediate) between the annelids and the arthropods. These mostly-tropical invertebrates, with only 75 extant species, live on the forest floor among moist, decaying leaves and feed on a mixed diet of plant and animal tissues. Like annelid worms, the Onychophorans have segmented bodies containing paired excretory organs (nephridia) and a combination of both male and female sex organs (monoecious). They lack a true exoskeleton, but the body is covered by a soft, chitinous cuticle. In true arthropod fashion, this cuticle is periodically shed (molted) to allow for continued growth. Onychophorans also share numerous other characteristics with the arthropods, including antennae, an open circulatory system, walking legs with claws, paired mandibles, and a system of slender air tubes (tracheae) for respiration.

Today, the prevailing opinion among modern taxonomists is that Onychophora and Arthropoda represent sister groups (see Cladogram 1), but the issue is far from settled. Both Anderson (1973) and Manton (1977) contend that onychophoran leg structure, locomotion, and embryological development is most comparable to that of the myriapods (millipedes, centipedes, et al.) and a recent comparison of nucleotide sequences in ribosomal RNA of velvet worms and other arthropods also supports a close affinity between these groups (Ballard et al. 1992).
Velvet worms may or may not prove to be direct ancestors of the arthropods, but regardless of classification, it seems reasonable to assume that the very first arthropods were distinctly worm-like in structure and appearance. They were also different in at least one important way: they had an exoskeleton. This structure, secreted by epidermal cells, was more rigid than annelid cuticle and necessitated membranous “joints” to provide the flexibility needed for movement. This exoskeleton (and its jointed appendages) is the hallmark of the arthropods.
Arthropod Evolution
All arthropods differ from their annelid ancestors in several important ways: they have a chitinous exoskeleton, jointed/segmented appendages, a well-developed head and mouthparts, striated muscles, and an open circulatory system with a dorsal heart. Unlike annelids, the arthropods produce large, yolk-laden eggs that are encased in a proteinaceous shell. Ciliated nephridia, paired segmental excretory organs found in annelids and onychophorans, have been replaced in arthropods by specialized excretory organs located on the head (green glands in crustacea), near the legs (coxal glands in horseshoe crabs), or in the abdomen (Malpighian tubules in terrestrial arthropods). Remarkable similarities in the chemical composition of the exoskeleton and in the ultrastructure of the compound eyes in organisms as diverse as millipedes, shrimp, and horseshoe crabs provide strong evidence that all of these groups must have evolved from a common ancestor.
The phylum Arthropoda, however, includes an incredibly diverse assortment of animals. They live in almost every type of habitat — from the deep oceans to hot deserts and from coastal estuaries to mountain peaks. Their ecological roles include everything from herbivores to vertebrate parasites, benthic scavengers to filter feeders, and zooplankton to pollinators of flowering plants. Phylogenomic analyses of nucleotide and amino acid sequences from a wide variety of these arthropods have provided strong support for the current hypothesis that they diverged into four distinct evolutionary lineages: Trilobita, Myriapoda, Chelicerata, and Pancrustacea. More about
Trilobites The trilobites became extinct by the end of the Permian Period and therefore represent an evolutionary “dead end”. Myriapods include millipedes, centipedes and their close relatives; they are mostly terrestrial animals with two body regions (a head and a trunk). The Chelicerata includes all arthropods with fang-like mouthparts (chelicerae): spiders, ticks, and mites (Arachnida), horseshoe crabs (Merostomata), and sea spiders (Pycnogonida). Pancrustacea includes all crustaceans (mostly marine) and all hexapods (mostly terrestrial and fresh-water).
Myriapoda
 The myriapods include both Chilopoda (centipedes) and Diplopoda (millipedes) as well as two other classes that are not as well-known: Symphyla and Pauropoda. All of these organisms live in moist environments near the soil surface (e.g., under stones, amid leaf mold, in rotting wood, etc.). Centipedes are primarily predators, the others are scavengers and herbivores.
The myriapods include both Chilopoda (centipedes) and Diplopoda (millipedes) as well as two other classes that are not as well-known: Symphyla and Pauropoda. All of these organisms live in moist environments near the soil surface (e.g., under stones, amid leaf mold, in rotting wood, etc.). Centipedes are primarily predators, the others are scavengers and herbivores.
 All myriapods exhibit ametabolous development (there is no significant change in body form as they mature). Eggs hatch into immatures (called young) that are similar to adults in most respects except size and sexual maturity. Like all other arthropods, they grow by molting, but they may also increase in length by adding an additional body segment at each molt. This type of growth (called anamorphosis) is common in myriapods, but it occurs only rarely in more “advanced” arthropods and is usually regarded as a “primitive” characteristic (pleisiomorphic condition).
All myriapods exhibit ametabolous development (there is no significant change in body form as they mature). Eggs hatch into immatures (called young) that are similar to adults in most respects except size and sexual maturity. Like all other arthropods, they grow by molting, but they may also increase in length by adding an additional body segment at each molt. This type of growth (called anamorphosis) is common in myriapods, but it occurs only rarely in more “advanced” arthropods and is usually regarded as a “primitive” characteristic (pleisiomorphic condition).
 Myriapods have only two functional body regions: a head and a trunk. The head is specialized for sensing the environment (eyes and antennae) and ingesting food. The trunk is adapted for locomotion and also contains most of the internal organs. Pauropods and symphylans are generally small (under 5 mm in length) with little or no body pigmentation — they are frequently creamy white in color. Symphylans have beaded antennae and 10 to 12 pairs of legs. Pauropods have branched antennae, nine pairs of legs, and plates of exoskeleton on the dorsal surface that usually span more than one body segment. Millipedes and centipedes are usually larger (more than 5 mm in length) and dark brown to black (although some species may be quite colorful). Centipedes have at least 15 pairs of legs — the first pair, immediately behind the head, contain poison glands and are modified as fangs for killing prey. Millipedes have at least 30 pairs of legs, two pair per body segment. They are not as fast-moving as centipedes and often curl into a ball or release a defensive spray as protection from predators.
Myriapods have only two functional body regions: a head and a trunk. The head is specialized for sensing the environment (eyes and antennae) and ingesting food. The trunk is adapted for locomotion and also contains most of the internal organs. Pauropods and symphylans are generally small (under 5 mm in length) with little or no body pigmentation — they are frequently creamy white in color. Symphylans have beaded antennae and 10 to 12 pairs of legs. Pauropods have branched antennae, nine pairs of legs, and plates of exoskeleton on the dorsal surface that usually span more than one body segment. Millipedes and centipedes are usually larger (more than 5 mm in length) and dark brown to black (although some species may be quite colorful). Centipedes have at least 15 pairs of legs — the first pair, immediately behind the head, contain poison glands and are modified as fangs for killing prey. Millipedes have at least 30 pairs of legs, two pair per body segment. They are not as fast-moving as centipedes and often curl into a ball or release a defensive spray as protection from predators.
Chelicerata
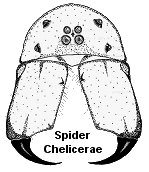 Chelicerate arthropods represent an ancient lineage of animals that are mostly adapted as predators of other organisms. They lack antennae but have six pairs of segmented appendages: fang-like chelicerae and pedipalps (which serve as mouthparts) and four pairs of walking legs. Chelicerae usually bear fangs and a poison gland for subduing prey. This subphylum is divided into three distinctive classes:
Chelicerate arthropods represent an ancient lineage of animals that are mostly adapted as predators of other organisms. They lack antennae but have six pairs of segmented appendages: fang-like chelicerae and pedipalps (which serve as mouthparts) and four pairs of walking legs. Chelicerae usually bear fangs and a poison gland for subduing prey. This subphylum is divided into three distinctive classes:
-
- Merostomata (horseshoe crabs) — Marine predators with a horseshoe-shaped body and abdominal book gills. Only four species remain from this ancient group of animals that are essentially unchanged from their ancestors that were common throughout the world’s oceans at least 200 million years ago.
- Pycnogonida (sea spiders) — Small, marine predators with thin bodies and long, slender legs. About 1000 species can be found throughout the world’s oceans but they are most commonly seen in the subtidal zones of polar waters.
- Arachnida (spiders, ticks, mites, scorpions, and their relatives) — These are mainly terrestrial predators with bodies that are usually divided into an anterior prosoma (=cephalothorax) and a posterior opisthosoma. In ticks and mites (order Acari) the prosoma and opisthosoma are broadly fused into a single body region.
Pancrustacea
The pancrustacean lineage contains a group of arthropods that appear to have diverged early in the Paleozoic era from a common ancestor with chewing mouthparts (mandibles). These include the crustaceans, the insects, and their close relatives. Most crustacea are found in marine habitats where they occupy a wide variety of ecological niches. Although a few species, such as pillbugs (also known as sowbugs or woodlice) live on land, they are restricted to moist habitats because they breathe with gills that function only when wet. Most crustacea have two body regions, a cephalothorax and an abdomen. All crustacea are distinctive from other arthropods because they have two pairs of antennae on the head and other appendages (including mouthparts and legs) that are biramous (two-branched).
 Based on recent phylogenomic evidence, hexapods appear to be an evolutionary offshoot of the Remipedia, a little-known class of crustaceans typically found in coastal cave systems (anchialine caves) that contain a mixture of fresh and saline water. Unlike remipedes, which superficially resemble myriapods, the hexapods have a body plan that is divided into three functional regions: head, thorax, and abdomen. The head bears compound eyes and a single pair of antennae, the thorax is especially adapted for locomotion with walking legs reduced to six in number, and the abdomen, which houses most of the internal organ systems, lacks segmental appendages except for the external genitalia.
Based on recent phylogenomic evidence, hexapods appear to be an evolutionary offshoot of the Remipedia, a little-known class of crustaceans typically found in coastal cave systems (anchialine caves) that contain a mixture of fresh and saline water. Unlike remipedes, which superficially resemble myriapods, the hexapods have a body plan that is divided into three functional regions: head, thorax, and abdomen. The head bears compound eyes and a single pair of antennae, the thorax is especially adapted for locomotion with walking legs reduced to six in number, and the abdomen, which houses most of the internal organ systems, lacks segmental appendages except for the external genitalia.
Cephalization was an important process in the evolutionary development of the hexapod lineage. As the head assumed a more prominent role in the organism’s survival, it increased in size and usefulness by assimilating adjacent trunk or body segments that previously had been adapted for locomotion. In this process, some leg appendages evolved into mouthparts that became adapted for catching, processing, or manipulating food items. These mouthparts are said to be serially homologous with walking legs.
Surviving in dry terrestrial environments also involved major changes in the respiratory system. Insects and other hexapods acquire oxygen directly from the air. An extensive system of tracheal tubes distributes air (containing oxygen) to every cell of the body. Desiccation is minimized by a series of valves (spiracles) that regulate the flow of air in and out of the body.

{kind=link}