
The Polyneopteran Orders
 Polyneopterans have a very simple, unspecialized body-plan that retains many of the ancestral (pleisiomorphic) characteristics of ametabolous insects: abdominal cerci, chewing mouthparts, long multi-segmented antennae, and a distributed nervous system with numerous segmental ganglia. Adults have four wings, although some species are secondarily wingless. The front wings (often called tegmina) are usually thickened or leathery. At rest, they cover and protect the hind wings. In flight, front and hind wings operate independently of one another (as in the Paleoptera). Hind wings are often enlarged near the base, providing a greater surface area for lift during flight. Most of the polyneopterans are rather weak or clumsy fliers.
Polyneopterans have a very simple, unspecialized body-plan that retains many of the ancestral (pleisiomorphic) characteristics of ametabolous insects: abdominal cerci, chewing mouthparts, long multi-segmented antennae, and a distributed nervous system with numerous segmental ganglia. Adults have four wings, although some species are secondarily wingless. The front wings (often called tegmina) are usually thickened or leathery. At rest, they cover and protect the hind wings. In flight, front and hind wings operate independently of one another (as in the Paleoptera). Hind wings are often enlarged near the base, providing a greater surface area for lift during flight. Most of the polyneopterans are rather weak or clumsy fliers.
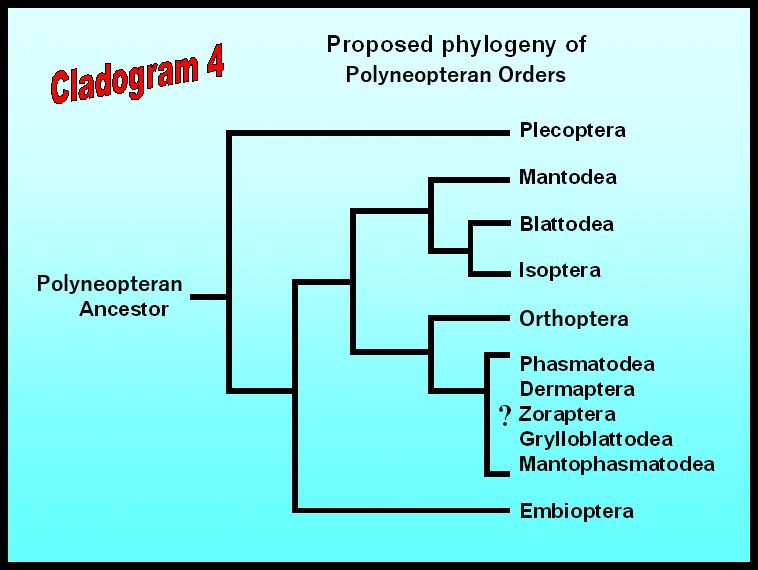
There is extensive controversy over phylogenetic relationships within the Polyneoptera complex. Although the fossil record contains many primitive neopterans, few systematists agree on how these extinct organisms are related to living orders and families. The ordinal status of modern-day polyneopterans is also the subject of much debate: “lumpers” are inclined to group all of these insects into five or six orders, whereas “splitters” divide them into as many as ten different orders.
Under the classification scheme we have chosen to use in this course (Cladogram 4), each major ecological group is given ordinal status. This may please the “splitters” but it probably gives a false impression that the evolutionary history of these organisms is more diverse than it really is. In fact, there is strong justification for combining some of these orders, and we will try to emphasize these groupings in the following paragraphs.
Plecoptera
 The first polyneopteran insects were scavengers and/or herbivores. From a physical standpoint, they were probably very similar to members of the present-day order Plecoptera. These insects, commonly known as stoneflies, are generally regarded as the earliest group of Neoptera. They probably represent an evolutionary “dead end” that diverged well over 300 million years ago. Immature stoneflies are aquatic nymphs (naiads). They usually live beneath stones in fast-moving, well-aerated water. Oxygen diffuses through the exoskeleton or into tracheal gills located on the thorax, behind the head, or around the anus.
The first polyneopteran insects were scavengers and/or herbivores. From a physical standpoint, they were probably very similar to members of the present-day order Plecoptera. These insects, commonly known as stoneflies, are generally regarded as the earliest group of Neoptera. They probably represent an evolutionary “dead end” that diverged well over 300 million years ago. Immature stoneflies are aquatic nymphs (naiads). They usually live beneath stones in fast-moving, well-aerated water. Oxygen diffuses through the exoskeleton or into tracheal gills located on the thorax, behind the head, or around the anus.
More about
Plecoptera Most species feed on algae and other submerged vegetation, but two families (Perlidae and Chloroperlidae) are predators of mayfly nymphs (Ephemeroptera) and other small aquatic insects. Adult stoneflies are generally found on the banks of streams and rivers from which they have emerged. They are not active fliers and usually remain near the ground where they feed on algae or lichens. In many species, the adults are short-lived and do not have functional mouthparts. Stoneflies are most abundant in cool, temperate climates.
Embioptera
 The order Embioptera (webspinners or embiids) is another group within the Polyneoptera complex that probably appeared early in the Carboniferous period. Many insect taxonomists believe webspinners may represent another evolutionary “dead end” that diverged about the same time as Plecoptera. Determining phylogenetic relationships for this group is unusually difficult because the Embioptera have a number of adaptations not found in any other insects. The tarsi of the front legs, for example, are enlarged and contain glands that produce silk. No other group of insects, fossil or modern, have silk-producing glands in the legs. The silk is used to construct elaborate nests and tunnels under leaves or bark. Webspinners live gregariously within these silken nests, feeding on grass, dead leaves, moss, lichens, or bark. Nymphs and adults are similar in appearance. Embiids rarely leave their silken tunnels; a colony grows by expanding its tunnel system to new food resources. Well-developed muscles in the hind legs allow these insects to run backward through their tunnels as easily as they run forward. Only adult males have wings. Front and hind wings are similar in shape and unusually flexible; they fold over the head when the insect runs backward through its tunnels.
The order Embioptera (webspinners or embiids) is another group within the Polyneoptera complex that probably appeared early in the Carboniferous period. Many insect taxonomists believe webspinners may represent another evolutionary “dead end” that diverged about the same time as Plecoptera. Determining phylogenetic relationships for this group is unusually difficult because the Embioptera have a number of adaptations not found in any other insects. The tarsi of the front legs, for example, are enlarged and contain glands that produce silk. No other group of insects, fossil or modern, have silk-producing glands in the legs. The silk is used to construct elaborate nests and tunnels under leaves or bark. Webspinners live gregariously within these silken nests, feeding on grass, dead leaves, moss, lichens, or bark. Nymphs and adults are similar in appearance. Embiids rarely leave their silken tunnels; a colony grows by expanding its tunnel system to new food resources. Well-developed muscles in the hind legs allow these insects to run backward through their tunnels as easily as they run forward. Only adult males have wings. Front and hind wings are similar in shape and unusually flexible; they fold over the head when the insect runs backward through its tunnels.
More about
Embioptera
Blood (hemolymph) is pumped into anterior veins to stiffen the wings during flight. In Embioptera, the mouthparts are directed forward (prognathous) rather than downward as in other primitive polyneopterans. This may simply be an adaptation for life in a tunnel, or as some taxonomists have suggested, it may mean that Embioptera are really more closely related to earwigs (order Dermaptera). Most Embioptera are tropical or subtropical.
Blattodea
 The ancestral prototype for the main line of Polyneoptera evolution was probably an insect very similar in appearance to a cockroach. Paleobiologists refer to this ancestral lineage as the Protoblattodean line. It probably dates from the early Carboniferous period, around 360 million years ago. In fact, fossil cockroaches found in late Carboniferous rock are remarkably similar to species living today.
The ancestral prototype for the main line of Polyneoptera evolution was probably an insect very similar in appearance to a cockroach. Paleobiologists refer to this ancestral lineage as the Protoblattodean line. It probably dates from the early Carboniferous period, around 360 million years ago. In fact, fossil cockroaches found in late Carboniferous rock are remarkably similar to species living today.
More about
Blattodea
In our scheme of classification, all modern cockroaches are grouped in one order, the Blattodea (or Blattaria). “Lumpers” often put them together with praying mantids (in the order Dictyoptera) or include them as a suborder of Orthoptera. The cockroaches, often known as “waterbugs” are scavengers or omnivores. They are most abundant in tropical or subtropical climates, but they also inhabit temperate and boreal regions. They are commonly found in close association with human dwellings where they are considered pests. Cockroaches have an oval, somewhat flattened body that is well-adapted for running and squeezing into narrow openings. Rather than flying to escape danger, roaches usually scurry into cracks or crevices. Much of the head and thorax is covered and protected dorsally by a large plate of exoskeleton (the pronotum).
When cockroaches lay eggs, the female’s reproductive system secretes a special capsule around her eggs. This structure, known as an öotheca, may be dropped on the ground, glued to a substrate, or retained within the female’s body. Production of an öotheca is a special adaptation found only in the cockroaches and praying mantids. This similarity suggests a close phylogenetic relationship between these groups and explains why some taxonomists prefer to lump them into a single order (Dictyoptera).
Mantodea
 From an ecological standpoint, cockroaches and mantids could not be more different: More about
From an ecological standpoint, cockroaches and mantids could not be more different: More about
Mantodea roaches are nocturnal scavengers, mantids are diurnal predators — in fact, they are the largest group of predators in the entire Polyneoptera complex. Mantids, order Mantodea, have elongate bodies that are specialized for a predatory lifestyle: long front legs with spines for catching and holding prey, a head that can turn from side to side, and cryptic coloration for hiding in foliage or flowers. Mantids are most abundant and most diverse in the tropics; there are only 5 species commonly collected in the United States and 3 of these have been imported from abroad.
Isoptera
 The termites, order Isoptera, are another group of insects that appear to be closely related to cockroaches. This conclusion is based on behavioral and ecological similarities between termites and wood roaches (members of the family Cryptocercidae). These cockroaches live in fallen timber on the forest floor, feeding on wood fibers which are then digested by symbiotic microorganisms within their digestive systems. They live in small family groups where each female provides care for her young offspring. Termites and wood roaches are thought to be close relatives because they both occupy similar habitats, share the same type of food resources, have the same intestinal symbionts, and provide care for their offspring.
The termites, order Isoptera, are another group of insects that appear to be closely related to cockroaches. This conclusion is based on behavioral and ecological similarities between termites and wood roaches (members of the family Cryptocercidae). These cockroaches live in fallen timber on the forest floor, feeding on wood fibers which are then digested by symbiotic microorganisms within their digestive systems. They live in small family groups where each female provides care for her young offspring. Termites and wood roaches are thought to be close relatives because they both occupy similar habitats, share the same type of food resources, have the same intestinal symbionts, and provide care for their offspring.
Termites are the only hemimetabolous insects that exhibit true social behavior. More about
Isoptera They build large communal nests that house an entire colony. Each nest contains adult reproductives (one queen and one king) plus hundreds or thousands of immatures that serve as workers and soldiers. Like cockroaches and mantids, the termites are most abundant in tropical and subtropical climates.
In Blattodea, Mantodea, and Isoptera, wing movement (particularly the downstroke) is largely dependent on muscles attached to the base of the wing (direct flight muscles). But in another branch of the Protoblattodean lineage, direct flight muscles are smaller and more of the power for flight is provided by indirect flight muscles (located in the thorax but not attached directly to the wings). At least two extinct orders (Protorthoptera and Protelytroptera) appear to be part of this second branch which also includes all the rest of the modern-day Polyneoptera orders: Orthoptera, Phasmatodea, Dermaptera, Grylloblattodea, and perhaps Zoraptera and Mantophasmatodea.
Orthoptera
 Orthoptera (grasshoppers, crickets, and katydids) probably arose during the middle of the Carboniferous period. Most living members of this order are terrestrial herbivores with modified hind legs that are adapted for jumping. Slender, thickened front wings (tegmina) fold back over the abdomen to protect membranous, fan-shaped hind wings. Many species have the ability to make and detect sounds. Orthoptera is one of the largest and most important groups of plant-feeding insects.
Orthoptera (grasshoppers, crickets, and katydids) probably arose during the middle of the Carboniferous period. Most living members of this order are terrestrial herbivores with modified hind legs that are adapted for jumping. Slender, thickened front wings (tegmina) fold back over the abdomen to protect membranous, fan-shaped hind wings. Many species have the ability to make and detect sounds. Orthoptera is one of the largest and most important groups of plant-feeding insects.
More about
Orthoptera
Although their phylogeny is not clear, all other members of the Polyneoptera complex are probably sister groups to the Orthoptera. These include the earwigs (order Dermaptera), leaf and stick insects (order Phasmatodea), rock crawlers (order Grylloblattodea), gladiators (order Mantophasmatodea), and zorapterans (order Zoraptera). Nearly all of these insects are herbivores or scavengers. In earwigs and stick insects, the chewing mouthparts are directed forward (prognathous) as in Embioptera; in rock crawlers, gladiators, and zorapterans, the mouthparts are directed downward (hypognathous) as in all other polyneopterans.
Phasmatodea
 The leaf and stick insects (order Phasmatodea or Phasmida) are sometimes grouped as a family or suborder of Orthoptera. All species are herbivores. As the name “walkingstick” implies, most phasmids are slender, cylindrical, and cryptically colored to resemble the twigs and branches on which they live. Members of the family Timemidae (=Phyllidae) bear a strong resemblance to leaves: abdomens are broad and flat, legs have large lateral extensions, and coloration is primarily brown, green, or yellow. More about
The leaf and stick insects (order Phasmatodea or Phasmida) are sometimes grouped as a family or suborder of Orthoptera. All species are herbivores. As the name “walkingstick” implies, most phasmids are slender, cylindrical, and cryptically colored to resemble the twigs and branches on which they live. Members of the family Timemidae (=Phyllidae) bear a strong resemblance to leaves: abdomens are broad and flat, legs have large lateral extensions, and coloration is primarily brown, green, or yellow. More about
Phasmatodea Most walkingsticks are slow-moving insects, a behavior pattern that is consistent with their cryptic lifestyle. In a few tropical species, the adults have well-developed wings, but most phasmids are brachypterous (reduced wings) or secondarily wingless.
Stick insects are most abundant in the tropics where some species may grow to 30 cm (12 inches) in length. Females do not have a well-developed ovipositor so they cannot insert their eggs into host plant tissue like most other Orthoptera. Instead, the eggs are dropped singly to the ground, sometimes from great heights.
Dermaptera
 Earwigs (order Dermaptera) are mostly scavengers or herbivores that hide in dark recesses during the day and become active at night. They feed on a wide variety of plant or animal matter. A few species may be predatory. Females lay their eggs in the soil and may guard them until they hatch. In a few species, maternal care even extends through the first two instars. Nymphs are similar in appearance to adults, but lack wings. The front wings are short, thick, and serve as protective covers for the hind wings. Hind wings are large, fan-shaped and pleated. They fold (both length-wise and cross-wise) to fit beneath the front wings when not in use. Some species are secondarily wingless. More about
Earwigs (order Dermaptera) are mostly scavengers or herbivores that hide in dark recesses during the day and become active at night. They feed on a wide variety of plant or animal matter. A few species may be predatory. Females lay their eggs in the soil and may guard them until they hatch. In a few species, maternal care even extends through the first two instars. Nymphs are similar in appearance to adults, but lack wings. The front wings are short, thick, and serve as protective covers for the hind wings. Hind wings are large, fan-shaped and pleated. They fold (both length-wise and cross-wise) to fit beneath the front wings when not in use. Some species are secondarily wingless. More about
Dermaptera In most earwigs, the cerci at the end of the abdomen are enlarged and thickened to form pincers (forceps). These pincers are used in grooming, defense, courtship, and even to help fold the hind wings.
The Dermaptera contains three suborders. Most species belong to the Forficulina. The other two groups, Arixeniina and Hemimerina, live in close association with mammals. The former (five species) live on Asian bats and the latter (eleven species) live on African rodents. All of these insects are adapted for a parasitic or semi-parasitic lifestyle: they are secondarily wingless and the cerci are not well-developed into pincers. Members of the Arixeniina give birth to live nymphs (vivipary).
Grylloblattodea
 The rock crawlers (order Grylloblattodea) are a small and obscure group of insects found only at high elevations in the mountains of China, Siberia, Japan, and western United States and Canada. Cave-dwelling species have been found in Korea and Japan. More about
The rock crawlers (order Grylloblattodea) are a small and obscure group of insects found only at high elevations in the mountains of China, Siberia, Japan, and western United States and Canada. Cave-dwelling species have been found in Korea and Japan. More about
Grylloblattodea These omnivorous insects scavenge for food on the surface of snowfields, under rocks, or near melting ice. They are active only at cold temperatures and move downward toward permafrost during warm seasons. As their name implies, rock crawlers have a blend of physical characteristics from both crickets (gryllo-) and cockroaches (blatta-). Some taxonomists include these insects as a suborder or family within Orthoptera. Others believe these insects are the only survivors of a primitive lineage that gave rise to other polyneopteran orders.
Mantophasmatodea
 The order Mantophasmatodea includes a very small group of insects that were first recognized as a separate order in 2002. More about
The order Mantophasmatodea includes a very small group of insects that were first recognized as a separate order in 2002. More about
Mantophasmatodea So far, living members of this group have been found only in the Brandberg and Erongo Mountains of Namibia and the Western Cape Province of South Africa. These insects appear to be nocturnal predators. They live within rock crevices, hide in clumps of grass, and prey on spiders and other small insects. As their order name suggests, they seem to exhibit a blend of the physical and ecological characteristics found in praying mantids (Mantodea) and walkingsticks (Phasmatodea).
Zoraptera
 Zoraptera, the final order within the Polyneoptera complex, is probably the most controversial in terms of its phylogenetic position within the class Insecta. In many respects, the Zoraptera are typical polyneopterans: they have chewing mouthparts, unsegmented cerci, and a striking resemblance to termites. But other features are more typical of insects in the Paraneoptera complex: the front wings (when present) are larger than the hind wings and have reduced venation, the nervous system has a reduced number of abdominal ganglia, and there are very few Malpighian tubules (excretory structures) in the digestive system.
Zoraptera, the final order within the Polyneoptera complex, is probably the most controversial in terms of its phylogenetic position within the class Insecta. In many respects, the Zoraptera are typical polyneopterans: they have chewing mouthparts, unsegmented cerci, and a striking resemblance to termites. But other features are more typical of insects in the Paraneoptera complex: the front wings (when present) are larger than the hind wings and have reduced venation, the nervous system has a reduced number of abdominal ganglia, and there are very few Malpighian tubules (excretory structures) in the digestive system.
This blend of Polyneopteran and Paraneoteran characteristics has led some entomologists to propose that Zoraptera represent a link between the two evolutionary lineages. Others reject this idea and claim that Zoraptera should be grouped with the protoblattodean lineage, near cockroaches, termites, and mantids. Still others argue that Zoraptera is a descendant of the protelytropteran lineage and therefore related to Dermaptera and Grylloblattodea. Regardless of phylogenetic placement, it seems likely that some of Zoraptera’s derived (apomorphic) characteristics are the result of convergent evolution.
More about
Zoraptera
Members of the order Zoraptera are small (less than 4 mm) and usually found in rotting wood, under bark, or in piles of old sawdust. They live in small aggregations and appear to scavenge on spores and mycelium of fungi, or occasionally, on mites and other small arthropods. Little more is known about their biology. Some Zoraptera are blind, pale in color, and wingless, while other members of the same species may be darkly pigmented with compound eyes and wings. The winged individuals are rather uncommon; they may be dispersal forms. The wings break off easily near the base, leaving only stubs.

{kind=link}