
Innate Behavior
Innate behavior is genetically programmed. Individuals inherit a suite of behaviors (often called an ethogram) just as they inherit physical traits such as body color and wing venation. In general, innate behaviors will always be:
- Heritable — encoded in DNA and passed from generation to generation
- Intrinsic — present in animals raised in isolation from others
- Stereotypic — performed in the same way each time by each individual
- Inflexible — not modified by development or experience
- Consummate — fully developed or expressed at first performance
Since innate behavior is encoded in DNA, it is subject to genetic change through mutation, recombination, and natural selection. Just like physical traits, innate behaviors are phylogenetic adaptations that have an evolutionary history.
 Comparative study of similar species often sheds light on the selective pressures that drive evolutionary changes in behavior. It may also help explain the origin of some very unusual behavior. One species of dance fly, for example, has a courtship ritual in which a male gives a ball of silk to a female. She unravels the ball while he mates with her. By itself, this curious behavior seems truly bizarre. But a study of courtship in other dance flies reveals that males use a nuptial gift as a way to divert a female’s aggressive behavior long enough for insemination to occur. In “primitive” species, the nuptial gift is an item of prey that the female consumes during copulation. In more “advanced” species, males wrap the prey in silk, thus buying a little extra time for copulation. In the species where males offer just a ball of silk, they are exploiting the female’s innate response to the stimulus of a nuptial gift. Just another example of “selfish genes” at work!
Comparative study of similar species often sheds light on the selective pressures that drive evolutionary changes in behavior. It may also help explain the origin of some very unusual behavior. One species of dance fly, for example, has a courtship ritual in which a male gives a ball of silk to a female. She unravels the ball while he mates with her. By itself, this curious behavior seems truly bizarre. But a study of courtship in other dance flies reveals that males use a nuptial gift as a way to divert a female’s aggressive behavior long enough for insemination to occur. In “primitive” species, the nuptial gift is an item of prey that the female consumes during copulation. In more “advanced” species, males wrap the prey in silk, thus buying a little extra time for copulation. In the species where males offer just a ball of silk, they are exploiting the female’s innate response to the stimulus of a nuptial gift. Just another example of “selfish genes” at work!
Just because an insect’s behavior is innate does not necessarily mean it is simple. Over time, natural selection can lead to surprisingly intricate and sophisticated behavior such as the dance language of honey bees or the courtship rituals of dance flies. These behaviors may appear purposeful and intelligent, but they are merely the product of millions of years of genetic refinement through natural selection.
In general, innate behaviors are viewed as “programmed” responses to external stimuli. They usually fit into one of the following categories:
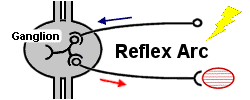 Reflex. The most basic unit of innate behavior is a simple reflex arc. This is a neural pathway that may involve as few as two neurons: a sensory neuron detects a stimulus and is linked with a motor neuron that sets off a response in an effector cell (such as a muscle or a gland cell). More commonly, reflex arcs also include an association neuron spliced between the sensory and motor neurons. The association neuron also synapses with other neurons to relay information to the brain and other parts of the body. Most insects have simple “startle” reflexes triggered by small disturbances as well as more comprehensive “escape” reflexes triggered by larger disturbances.
Reflex. The most basic unit of innate behavior is a simple reflex arc. This is a neural pathway that may involve as few as two neurons: a sensory neuron detects a stimulus and is linked with a motor neuron that sets off a response in an effector cell (such as a muscle or a gland cell). More commonly, reflex arcs also include an association neuron spliced between the sensory and motor neurons. The association neuron also synapses with other neurons to relay information to the brain and other parts of the body. Most insects have simple “startle” reflexes triggered by small disturbances as well as more comprehensive “escape” reflexes triggered by larger disturbances.
Orientation Behaviors are coordinated movements (walking, flying, swimming, etc.) that occur in response to an external stimulus. These behaviors have adaptive value for survival by helping the insect locate (or avoid) the source of a stimulus. Orientation behaviors can be viewed as elements in a neural hierarchy. The simplest behaviors involve input from only a single sensory receptor whereas more advanced behaviors require bilateral input from a pair of receptors.
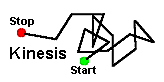 Kinesis is a change in the speed of movement (orthokinesis) or a change in the rate of turning (klinokinesis) which is directly proportional to the intensity of a stimulus. Input from only a single sensory receptor is necessary. A kinesis is non-directed orientation, that is, the animal exhibits a “random walk”. The change in speed or rate of turning increases the probability of locating the stimulus but does not guarantee it.
Kinesis is a change in the speed of movement (orthokinesis) or a change in the rate of turning (klinokinesis) which is directly proportional to the intensity of a stimulus. Input from only a single sensory receptor is necessary. A kinesis is non-directed orientation, that is, the animal exhibits a “random walk”. The change in speed or rate of turning increases the probability of locating the stimulus but does not guarantee it.
Taxis is a movement directly toward (positive) or away from (negative) a stimulus. A klinotaxis involves side-to-side motions of the head or body with successive comparison of stimulus intensity as the animal moves forward. A tropotaxis requires bilateral input from paired sensory receptors such that the signal is equalized in both receptors. Stimulus intensity increases with movement toward the source and decreases with movement away from the source. A prefix may also be used to designate the type of stimulus involved (i.e. phototaxis=light; geotaxis=gravity; thigmotaxis=contact with other objects
 The dorsal light reaction is a special case (telotaxis) in which movement occurs at a constant 90° angle to a light source. By keeping the sun directly overhead, a flying or swimming insect can insure that it travels parallel to the ground (or water surface). A diving beetle, for example, can be fooled into swimming upside down in an aquarium that is lit from below. The dorsal light reaction also explains why moths tend to circle a street lamp at night.
The dorsal light reaction is a special case (telotaxis) in which movement occurs at a constant 90° angle to a light source. By keeping the sun directly overhead, a flying or swimming insect can insure that it travels parallel to the ground (or water surface). A diving beetle, for example, can be fooled into swimming upside down in an aquarium that is lit from below. The dorsal light reaction also explains why moths tend to circle a street lamp at night.
The light compass reaction is another special case (menotaxis) in which insects (honey bees, for example) fly away from the nest site at a fixed angle (x°) to the sun and return at the supplementary angle (180-x°) — all without ever taking a class in geometry!
Fixed Action Pattern (FAP) is a sequence of coordinated movements that are performed together as a “unit” without interruption. Each FAP is triggered by a unique stimulus variously known as a sign stimulus, a key stimulus, or a releaser. A praying mantis striking at prey is a typical example. The releaser for this FAP is any movement by a small (prey-sized) object within striking distance. Once initiated, the mantis cannot change direction in mid-strike or abort the mission if the prey escapes. Other common examples of FAPs include courtship displays, hunting or food gathering, nest-building activities, and attack or escape movements. Unlike simple reflexes, FAPs may involve a whole-body response and often require a threshold level of internal readiness (drive).
A fixed action pattern is rarely triggered by the “big picture” (Gestalt) in an environmental context. Instead, the sign stimulus is usually a highly specific signal that is consistently encountered at an appropriate time. Thus a male fruit fly will perform a courtship display for a pheromone-impregnated cork even though the cork doesn’t look, taste, feel, or act like a female fruit fly. Case Study:
Sand Wasps and FAPs Sometimes, it is possible to find or create a supernormal stimulus, essentially an exaggerated signal that produces a more vigorous or more sustained response than a normal releaser. Apple growers, for example, hang large red spheres coated with stick-um in their orchards to catch adults of the apple maggot (Rhagoletis pomonella). The red spheres are a key stimulus for oviposition, and to female apple maggots, size matters!
Performing one FAP may lead an insect to encounter the releaser for a second FAP and that in turn may lead to the releaser for a third FAP, etc. This type of behavioral cascade is common in insects. Niko Tinbergen, one of the “fathers” of modern ethology, demonstrated that hunting behavior in a predatory wasp proceeds through a stepwise series of three FAPs. The first releaser is visual: movement of a prey-sized object triggers down-wind pursuit of the prey. At that point, prey odor is a releaser that triggers catching behavior. Finally, tactile cues from the prey release stinging and egg laying behavior.

When a fixed action pattern fulfills or satisfies a physiological drive, it may be known as a consummatory act. Taking a blood meal, for example, satisfies a mosquito’s hunger drive. Any behavior that increases an individual’s probability of encountering the releaser for a consummatory act is often called an appetative behavior. Thus, chirping by a cricket may be regarded as an appetative behavior because it increases the chances of finding a mate and satisfying the sex drive.
