Sex Determination
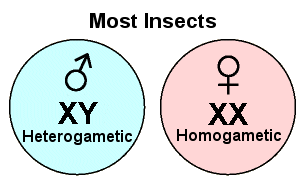 Like humans, most insects have a single pair of chromosomes that carry the genetic information for determining an individual’s gender. If an embryo inherits a pair of “X” chromosomes, it will develop as a female; if it inherits one “X” and one “Y”, it will develop as a male. The “XX” female is said to be homogametic; the “XY” male is heterogametic. In this case (as in humans) the male’s contribution determines the offspring’s gender. Some insect species have no “Y” chromosome at all — males have just one “X”, and females have two. A similar condition is found in some parthenogenetic species of aphids in which “maleness” occurs through the loss (degeneration) of one chromosome during embryogenesis. In both cases, the males end up with an odd number of chromosomes (2n-1).
Like humans, most insects have a single pair of chromosomes that carry the genetic information for determining an individual’s gender. If an embryo inherits a pair of “X” chromosomes, it will develop as a female; if it inherits one “X” and one “Y”, it will develop as a male. The “XX” female is said to be homogametic; the “XY” male is heterogametic. In this case (as in humans) the male’s contribution determines the offspring’s gender. Some insect species have no “Y” chromosome at all — males have just one “X”, and females have two. A similar condition is found in some parthenogenetic species of aphids in which “maleness” occurs through the loss (degeneration) of one chromosome during embryogenesis. In both cases, the males end up with an odd number of chromosomes (2n-1).
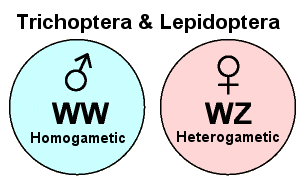 In Lepidoptera and Trichoptera, however, the homo- and heterogametic sexes are reversed: females are heterogametic and males are homogametic. To distinguish this system from standard X-Y sex determination, these sex chromosomes are designated “W” and “Z” (instead of “X” and “Y”). Thus, a female butterfly is “WZ” and a male butterfly is “WW”. In this case, the female’s contribution determines the offspring’s gender. Oddly, there is only one other group of organisms in the animal kingdom that has this pattern of sex determination. Can you name it?
In Lepidoptera and Trichoptera, however, the homo- and heterogametic sexes are reversed: females are heterogametic and males are homogametic. To distinguish this system from standard X-Y sex determination, these sex chromosomes are designated “W” and “Z” (instead of “X” and “Y”). Thus, a female butterfly is “WZ” and a male butterfly is “WW”. In this case, the female’s contribution determines the offspring’s gender. Oddly, there is only one other group of organisms in the animal kingdom that has this pattern of sex determination. Can you name it?
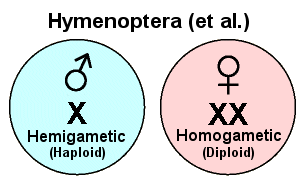 A third method of sex determination, called haplo-diploidy, is found in all Hymenoptera, many Thysanoptera, some scale insects (Hemiptera/Homoptera), and a few weevils (Coleoptera). These insects have diploid, homogametic females (“XX”), but all of the males are haploid — they develop by parthenogenesis (asexually) from unfertilized eggs. Primary oocytes undergo meiosis to form haploid eggs, but meiosis is unnecessary in primary spermatocytes because the cells are already haploid. Unmated females can lay eggs that will develop into males. Once a female mates and receives sperm from a male, she has two options:
A third method of sex determination, called haplo-diploidy, is found in all Hymenoptera, many Thysanoptera, some scale insects (Hemiptera/Homoptera), and a few weevils (Coleoptera). These insects have diploid, homogametic females (“XX”), but all of the males are haploid — they develop by parthenogenesis (asexually) from unfertilized eggs. Primary oocytes undergo meiosis to form haploid eggs, but meiosis is unnecessary in primary spermatocytes because the cells are already haploid. Unmated females can lay eggs that will develop into males. Once a female mates and receives sperm from a male, she has two options:
- She can produce a female offspring by opening the valve at the base of her spermatheca to release sperm onto the egg as it passes through her oviduct, or
- She can produce a male offspring by closing the spermathecal valve and preventing any sperm from reaching the egg.
Control over the gender of offspring has proven to be a useful adaptation for some insects. A biased sex ratio that favors females over males can reduce competition for limited food resources and increase the reproductive potential of the population. Bees, wasps, and ants form large colonies of queens and workers (all female) in which males are produced only sporadically as needed for reproduction.

 Today, all of the more “advanced” insects exhibit internal fertilization — males deposit their sperm inside a female’s body during an act of copulation. This novel adaptation, which appeared soon after insects diverged from their myriapod-like ancestors, presumably ensured that more sperm found their way to a receptive female. But the genetic programming for spermatophore production still persists in most modern insects. After a male deposits his spermatophore inside a female’s reproductive system, she digests the lipo-protein coat and uses it as a source of additional nutrition for her eggs. In some cases, the quality (or quantity) of this nuptial gift may even determine whether a female accepts or rejects the male’s gametes.
Today, all of the more “advanced” insects exhibit internal fertilization — males deposit their sperm inside a female’s body during an act of copulation. This novel adaptation, which appeared soon after insects diverged from their myriapod-like ancestors, presumably ensured that more sperm found their way to a receptive female. But the genetic programming for spermatophore production still persists in most modern insects. After a male deposits his spermatophore inside a female’s reproductive system, she digests the lipo-protein coat and uses it as a source of additional nutrition for her eggs. In some cases, the quality (or quantity) of this nuptial gift may even determine whether a female accepts or rejects the male’s gametes.